веществ на свету фотоавтотрофами при участии фотосинтетических пигментов. В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Фотосинтез – процесс преобразования энергии света в энергию АТФ презентация
Содержание
- 2. Фотосинтез - процесс преобразования энергии света в
- 3. Фотосинтез – процесс преобразования энергии света в
- 4. Зеленый свет хлорофилл пропускает. А остальные усваивает
- 6. Если возбудить изолированную молекулу хлорофилла, она флюоресциирует
- 7. Фотосинтез придумали сине-зеленые водоросли, они же цианобактерии Все остальные их просто инкорпорировали
- 9. F Фотосинтез – усвоение энергии света. Световая фаза фотосинтеза
- 11. Фотон ударяет в пигментную молекулу Фотосистемы
- 13. Фотосистема II - первый функциональный комплекс электрон-транспортной
- 15. Циклическое фосфорилирование обеспечивает синтез АТФ
- 19. А теперь синтез АТФ За счет градиента Концентрации протонов
- 20. Электрон-транспортная цепь расположена во внутренней мембране митохондрий
- 22. Рибулозобисфосфаткарбоксилаза (RuBisCO) — фермент, катализирующий присоединение углекислого

Слайд 3Фотосинтез – процесс преобразования энергии света в энергию АТФ
Не любая длина
волны света может
Быть усвоена хлорофиллом
Быть усвоена хлорофиллом




Слайд 7Фотосинтез
придумали
сине-зеленые
водоросли,
они же
цианобактерии
Все остальные
их просто
инкорпорировали



Слайд 11Фотон ударяет в пигментную молекулу
Фотосистемы II, повышая энергетический
уровень одного
из электронов. Этот электрон
отдает свою энергию соседнему пигменту и его
электрон переходит на более высокий уровень.
Так продолжается до тех пор, пока он не достигнет пары молекул хлорофилла а P680 в
реакционном центре комплекса Фотосистемы II
отдает свою энергию соседнему пигменту и его
электрон переходит на более высокий уровень.
Так продолжается до тех пор, пока он не достигнет пары молекул хлорофилла а P680 в
реакционном центре комплекса Фотосистемы II
Этот электрон передается на первичный
Акцептор электронов.
Димер хлорофилла а, П680+, катализирует
Расщепление воды на два электрона, два
Протона и атом кислорода.

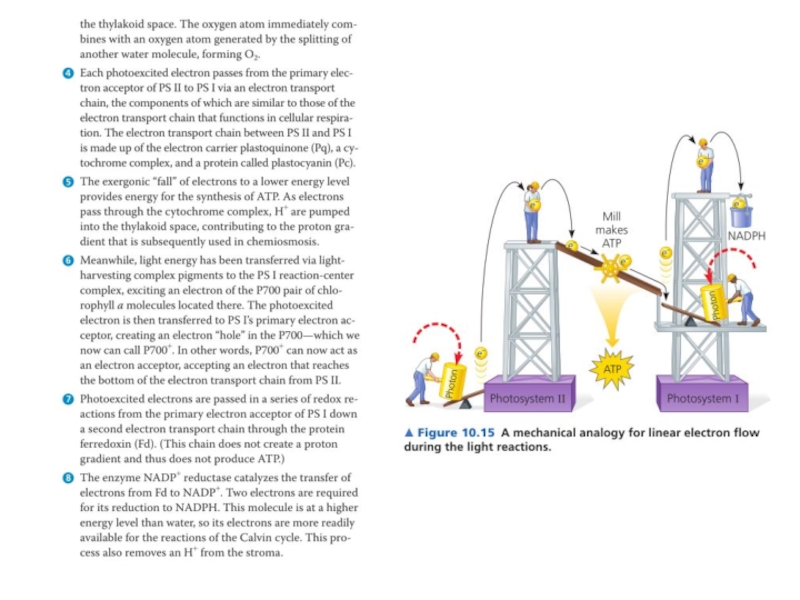
Слайд 13Фотосистема II - первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он
расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.
Окисляя воду, фотосистема II поставляет электроны в ЭТЦ хлоропласта, где они используются для восстановления НАДФ+ или циклического фосфорилирования. Помимо этого, окисление воды приводит к образованию протонов и формированию протонного градиента, используемого в дальнейшем для синтеза АТФ. Фотохимическое окисление воды, которое осуществляет фотосистема II, сопровождается выделением молекулярного кислорода. Этот процесс (составная часть фотосинтеза растений) является основным источником кислорода на Земле.
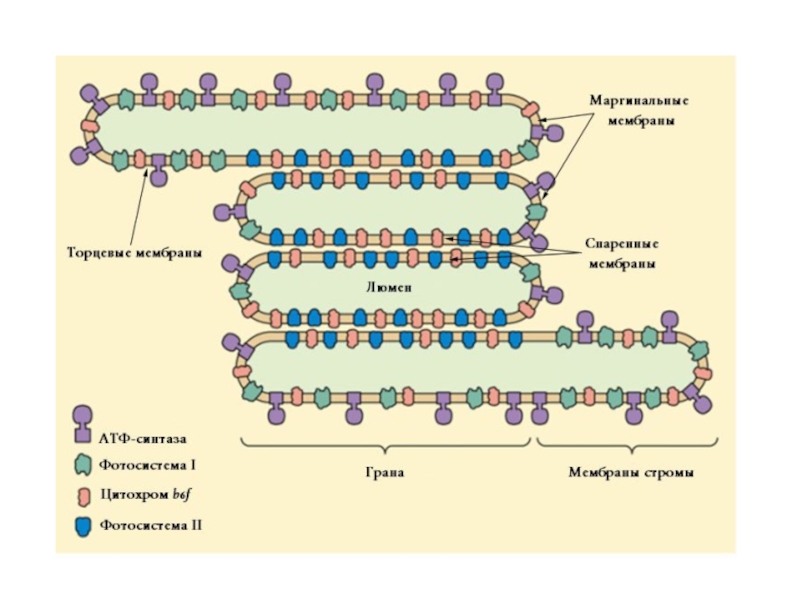
Основная функция фотосистемы II — генерация сильного окислителя, который индуцирует процесс окисление воды и перенос её электронов на мембранный переносчик[en]. Основная функция фотосистемы I — насытить эти низкоуровневые электроны энергией, чтобы с их помощью осуществить восстановление НАДФ+. Поскольку энергия суммарного процесса слишком велика, чтобы осуществить его в рамках одного реакционного центра, в ходе эволюции появились две фотосистемы, которые раздельно осуществляют разные части этой реакции. Их специфические функции и определяют особенности их строения. Так, фотосистема I симметрична, то есть в ней работают две ветви электронного транспорта, что делает его значительно более быстрым, в то время как фотосистема II асимметрична и обладает только одной рабочей ветвью, что замедляет транспорт электронов, но делает его более управляемым. Обе фотосистемы значительно отличаются по строению антенн, дополнительных субъединиц, способов регуляции и положению в мембране. Так, фотосистема I обладает интегральной антенной, хлорофиллы которой расположены непосредственно на главных белках комплекса — А и B, в то время как у фотосистемы II они вынесены на внешние белки CP47 и CP43. По количеству дополнительных малых регуляторных субъединиц ФСII значительно превосходит ФСI, что связано с необходимостью тонкой регуляции процесса окисления воды, который потенциально крайне опасен для клетки. Этим же объясняется неоднородное распределение фотосистем в мембране тилакоида: ФСI располагается преимущественно в области маргинальных, торцевых и стромальных мембран, а ФСII практически полностью находится в области спаренных мембран, что обеспечивает клетке дополнительную защиту от продуцируемых ею активных форм кислорода
Окисляя воду, фотосистема II поставляет электроны в ЭТЦ хлоропласта, где они используются для восстановления НАДФ+ или циклического фосфорилирования. Помимо этого, окисление воды приводит к образованию протонов и формированию протонного градиента, используемого в дальнейшем для синтеза АТФ. Фотохимическое окисление воды, которое осуществляет фотосистема II, сопровождается выделением молекулярного кислорода. Этот процесс (составная часть фотосинтеза растений) является основным источником кислорода на Земле.
Основная функция фотосистемы II — генерация сильного окислителя, который индуцирует процесс окисление воды и перенос её электронов на мембранный переносчик[en]. Основная функция фотосистемы I — насытить эти низкоуровневые электроны энергией, чтобы с их помощью осуществить восстановление НАДФ+. Поскольку энергия суммарного процесса слишком велика, чтобы осуществить его в рамках одного реакционного центра, в ходе эволюции появились две фотосистемы, которые раздельно осуществляют разные части этой реакции. Их специфические функции и определяют особенности их строения. Так, фотосистема I симметрична, то есть в ней работают две ветви электронного транспорта, что делает его значительно более быстрым, в то время как фотосистема II асимметрична и обладает только одной рабочей ветвью, что замедляет транспорт электронов, но делает его более управляемым. Обе фотосистемы значительно отличаются по строению антенн, дополнительных субъединиц, способов регуляции и положению в мембране. Так, фотосистема I обладает интегральной антенной, хлорофиллы которой расположены непосредственно на главных белках комплекса — А и B, в то время как у фотосистемы II они вынесены на внешние белки CP47 и CP43. По количеству дополнительных малых регуляторных субъединиц ФСII значительно превосходит ФСI, что связано с необходимостью тонкой регуляции процесса окисления воды, который потенциально крайне опасен для клетки. Этим же объясняется неоднородное распределение фотосистем в мембране тилакоида: ФСI располагается преимущественно в области маргинальных, торцевых и стромальных мембран, а ФСII практически полностью находится в области спаренных мембран, что обеспечивает клетке дополнительную защиту от продуцируемых ею активных форм кислорода
 хлоропластов. Он расположен в мембранах тилакоидов")

Слайд 16
Нециклический транспорт
электронов начинается с того, что марганцевый кластер фотосистемы II окисляет воду, восстанавливая пул пластохинонов. Далее цитохром b6f-комплекс окисляет пластохиноны, а электрон через пластоцианин передаётся на фотосистему I, где используется для синтеза НАДФН. Нарушение формальной логики в названиях фотосистем связано с тем, что фотосистема I была открыта раньше, чем фотосистема II.
Нециклический – синтез НАДН и АТФ, но АТФ мало




Слайд 20Электрон-транспортная цепь расположена во внутренней мембране
митохондрий и накачивает протоны в межмембранное
пространство
В хлоропластах этц накачивает протоны в пространство люмена тилакоидов,
Тоже межмембранное
В хлоропластах этц накачивает протоны в пространство люмена тилакоидов,
Тоже межмембранное


Слайд 22Рибулозобисфосфаткарбоксилаза (RuBisCO) — фермент, катализирующий присоединение углекислого газа к рибулозо-1,5-бифосфату на
первой стадии цикла Кальвина, а также реакцию окисления рибулозобифосфата на первой стадии процесса фотодыхания. Является одним из важнейших ферментов в природе, поскольку играет центральную роль в основном механизме поступления неорганического углерода в биологический круговорот. Рибулозобисфосфаткарбоксилаза является основным ферментом листьев растений и поэтому считается наиболее распространённым ферментом на Земле
У растений, цианобактерий и хемоавтотрофных
протеобактерий фермент обычно состоит
из двух типов белковых субъединиц:
Большая цепь (L, массой 55 000 Да)
Малая цепь (S, массой 13 000 Да)
Активный центр фермента находится на
больших цепях, объединённых в димеры.
Все 8 димеров больших цепей и 8 малых цепи
объединены в единый комплекс массой 540 000 Да.
У некоторых протеобактерий малых цепей в составе
рубиско не обнаружено. При этом большие цепи
кодируются в ДНК хлоропласта, а малые — в ядре
и транспортируются в хлоропласт перед сборкой всего белка.
Для работы фермента необходимы ионы Mg2+, которые размещаются в активном центре и способствуют присоединению CO2 к остатку лизина, в ходе чего образуется карбамат. Образование карбамата протекает легче в щелочной среде:
 — фермент, катализирующий присоединение углекислого газа к рибулозо-1,5-бифосфату на первой стадии цикла Кальвина,")