- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Физиология растений презентация
Содержание
- 1. Физиология растений
- 2. Механизмы поступления воды в растительную клетку.
- 3. Сильные (ковалентные) взаимодействия между молекулами воды обусловлены
- 4. Греческий алфавит:
- 5. В результате формирования большого числа водородных связей
- 6. Плотность воды в интервале температур от +4
- 7. Можно обобщить некоторые наиболее важные уникальные свойства
- 8. Диэлектрическая проницаемость воды равна 80,2 при 20
- 9. Притяжение между молекулами воды, наблюдаемое в жидкой
- 11. Вода в клетке находится в двух состояниях:
- 12. Гипертонический раствор Состояние протоплазмы зависит от
- 13. Гипертонический раствор Состояние протоплазмы зависит от активности
- 14. Гипертонический раствор Состояние протоплазмы зависит от
- 15. Как движется вода? Каков механизм? Почему так
- 16. эндоцитоз экзоцитоз Удивительно, но сейчас есть данные
- 17. Некоторые фундаментальные физические основы процессов движения воды
- 18. Химический потенциал воды μw выражает максимальное количество
- 19. - Активность чистой воды равна единице, а
- 20. Водный потенциал характеризует способность воды диффундировать, испаряться
- 21. - Водный потенциал чистой воды равен нулю.
- 22. Водный потенциал клетки, ткани или органа, целого
- 23. Матричный потенциал Ψm Связан со
- 24. Осмотический потенциал Ψр обусловлен наличием в клетках
- 25. Тургорный потенциал Ψt обусловлен эластичным противодействием клеточной
- 26. Поскольку в большинстве случаев величины как матричного,
- 27. Транспорт воды через мембраны клетки осуществляют специальные
- 28. Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины
- 29. Транспорт воды осуществляют специальные белки аквапорины, формирующие
- 30. MIPs - транспортируют Небольшие нейтральные молекулы – воду, глицерол, мочевину
- 31. снаружи цитоплазма Селективная к воде пора Одна
- 32. Аквапорины регулируются фосфорилированием как минимум трех
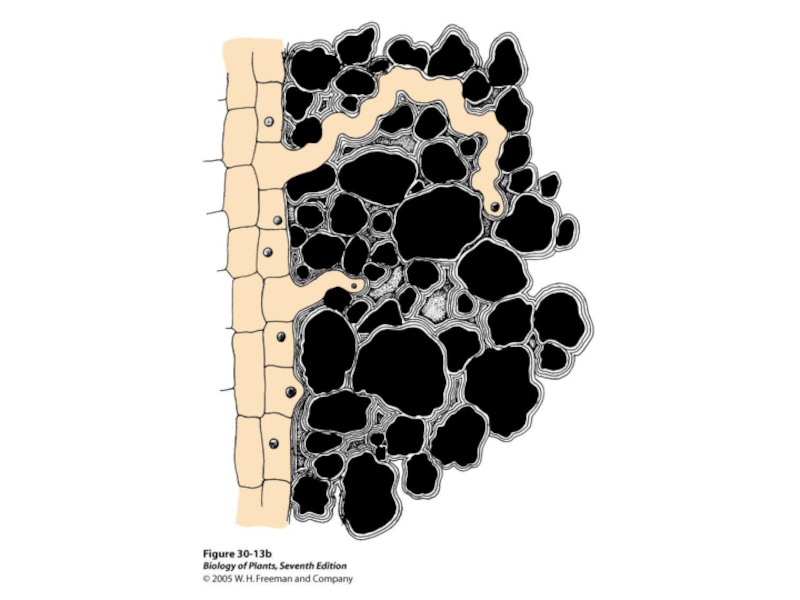
- 36. Корневые волоски
- 37. Корневые волоски
- 38. Апопласти-ческий путь Поясок Каспари
- 39. Корневые волоски арабидопсиса
- 40. Транспорт через плазматическую мембрану эпидермальных клеток корня
- 41. Эпидермальные клетки представлены (примерно 1:1) трихобластами (образующими
- 42. Листья (верхний концевой «двигатель») Градиент воды (направление
- 43. Гуттация, “слёзы растений”, это выделение воды из
- 44. воздух ψ = −100.0 Mпa
- 45. Замыкающие клетки устьиц – главные регуляторы траспирации
- 46. Перфориро- ванная пластинка Сосудистый элемент
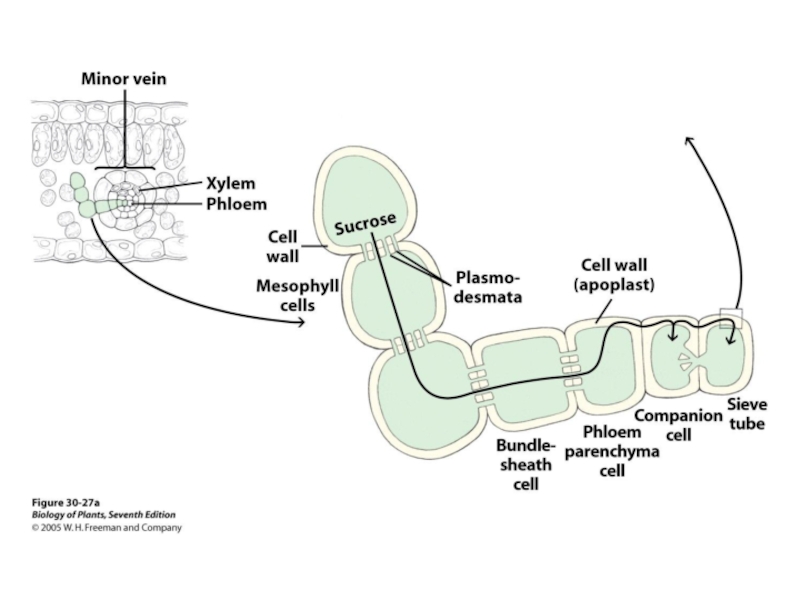
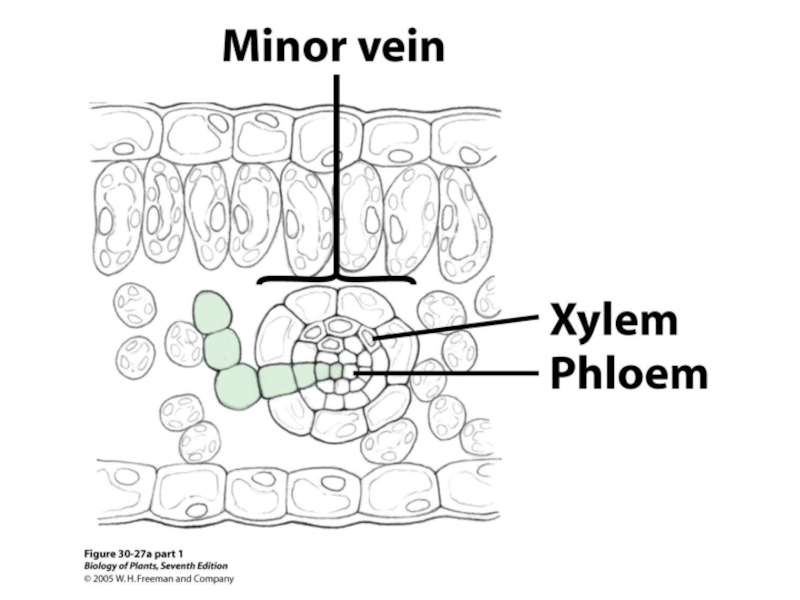
- 47. Флоэма Ситовидно-трубочный элемент (слева) и
- 59. Транспирация. Основным органом транспирации является
- 60. Транспирация. Основным органом транспирации является
- 62. В испарении принимают участие три структуры:
- 63. Транспирация. При устьичной транспирации выделяют
- 65. Интенсивность (скорость) транспирации определяется количеством граммов воды,
- 66. Г у т т а ц и
- 68. Системы, которые создают корневое давление и ответственны
- 69. Верхний концевой двигатель – системы, обеспечивающие присасывающую
- 70. Воздух Ψ = −100.0 Mpa Лист
- 71. 1 – вода обладает особой структурой молекулы,
- 72. Самостоятельно – по учебнику регуляция работы устьичного аппарата экзогенными и эндогенными факторами (глава 4).

Слайд 2
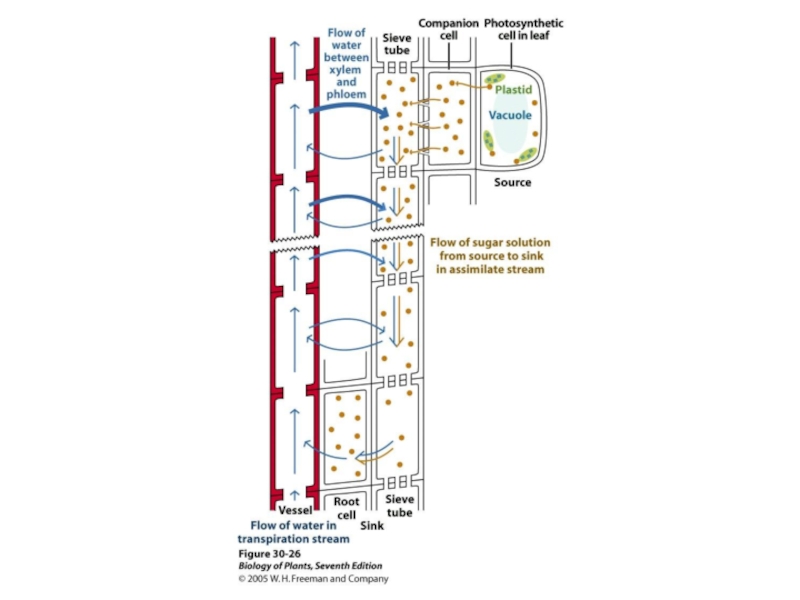
Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении.
План
- понятия осмоса, химического потенциала воды, водного потенциала клетки;
- строение и регуляция аквапоринов (белков. "катализирующих" транспорт воды в клетку)
- транспорт из почвы в корень, передвижение воды по растению, транспирация, роль устьиц, регуляция работы устьичного аппарата

Слайд 3Сильные (ковалентные) взаимодействия между молекулами воды обусловлены структурой молекул этого соединения.
Атом кислорода обладает сильной электроотрицательностью и стремится оттянуть электроны от атомов водорода. Благодаря этому на атоме кислорода возникает частично отрицательный заряд (δ–), в то время как два атома водорода приобретают положительный заряд (δ+).
Несущие положительный заряд атомы водорода испытывают электростатическое притяжение со стороны отрицательно заряженных атомов кислорода соседних молекул воды.
Водородная связь между молекулами воды имеет энергию 4,8 ккал/моль.
 взаимодействия между молекулами воды обусловлены структурой молекул этого соединения. Расстояние между ядром кислорода")

Слайд 5В результате формирования большого числа водородных связей возникает большая упорядоченность в
На отдельных участках жидкая вода приобретает почти кристаллическую структуру, что чрезвычайно важно, поскольку может играть определенную роль во взаимодействиях и ориентации молекул в водных растворах.
Вода обладает большой теплоемкостью. Удельной теплоемкостью воды называют количество теплоты, которое необходимо, чтобы поднять температуру 1 кг воды на 1о С.
Большая теплоемкость означает, что существенное увеличение тепловой энергии вызывает лишь сравнительно небольшое повышение ее температуры. Объясняется это во многом тем, что значительная часть энергии расходуется на разрыв водородных связей, ограничивающих подвижность молекул воды.

Слайд 6Плотность воды в интервале температур от +4 до 0 оС понижается,
Поскольку лед плавает в воде, он образуется при замерзании сначала на ее поверхности и лишь под конец в придонных слоях. Если бы замерзание шло в обратном порядке, то жизнь в пресноводных водоемах вообще не могла бы существовать.
Еще одной важной физической характеристикой воды является необычайно высокая диэлектрическая проницаемость (Д), что является следствием молекулярной структуры. Высокая диэлектрическая проницаемость воды делает электрические силы между растворенными в ней заряженными веществами относительно слабыми.

Слайд 7Можно обобщить некоторые наиболее важные уникальные свойства воды:
1 – термальные (точки
соединений О и Н), огромная теплоемкость, изменения формы – «легкая» твердая, тяжелая жидкая, газ.
2 – свойства растворителя (изменение точек плавления и кипения при растворении различных соединений), кислород и углекислота растворимы в воде
3 – особая проводимость для света (весь красный свет поглощается первым метром воды, весь желтый первыми 10 м, остальная высокоэнергетичная часть проходит в глубь, придавая голубой цвет океану)
4 – особенно высокая проводимость для звука

Слайд 8Диэлектрическая проницаемость воды равна 80,2 при 20 оС и 78,4 при
Для неполярной жидкости – гексана – Д = 1,87. Следовательно, электрическое притяжение для таких ионов, как Na+ и Cl–, в гексане больше (80,2/1,7) в 43 раза, чем в воде.
Значительно более сильное притяжение в гексане, чем в воде, уменьшает степень ионизации NaCl по сравнению с диссоциацией этой соли в водном растворе, т. е. вода – хороший растворитель для заряженных частиц.
Из всех жидкостей самое большое поверхностное натяжение у воды (поверхностное натяжение – результат действующих между молекулами сил на поверхности раздела фаз).

Слайд 9Притяжение между молекулами воды, наблюдаемое в жидкой фазе, обычно называют когезией,
В случае воды в стеклянных капиллярах или в сосудах ксилемы притяжение между молекулами воды и стенками велико, и поэтому жидкость поднимается.


Слайд 11Вода в клетке находится в двух состояниях: свободном и связанном (3-10
В вакуолях вода удерживается относительно низкомолекулярных соединений (осмотически связанная), и большая часть воды находится в свободном состоянии.
В клеточной стенке часть молекул воды адсорбируется на поверхности фибрилл клеточной стенки. Вода тут связывается, главным образом, целлюлозой, гемицеллюлозой, пектиновыми веществами, т. е. коллоидно-связанная вода.
Кроме того, в клеточной стенке есть свободная вода (в порах). В цитоплазме – свободная, коллоидно- и осмотически связанная вода.
Вода, которая находится на расстоянии 1 нм от поверхности белковой молекулы, связана сильно.
Осмотически связанная вода цитоплазмы – это вода, связанная с ионами.
,")
Слайд 12Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
Изотонический
раствор
Гипотонический
раствор
Плазмолиз
Нормальное
состояние
Тургесцентное
состояние
Вакуоль

Слайд 13Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
НОРМА:
Гипотонический
раствор
(ПОЧВА, АПОПЛАСТ)
клетки лука (внешний раствор: 20%
Опыт описан Архимедом.
клетки лука (внешний раствор: 20% сахарозы)Опыт описан Архимедом.")
Слайд 14Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
Изотонический
раствор
Гипотонический
раствор
Плазмолиз
Нормальное
состояние
Тургесцентное
состояние
Плазмолизованные клетки элодеи в растворе, содержащем 30% сахарозы - вода начинает выходить вгновенно
10 мкм

Слайд 15Как движется вода? Каков механизм? Почему так быстро?
Какой белок/ген регулирует
Этим занимается раздел Физиологии Растений называемый "Водообменом"
Н2О
Основной механизм траспорта воды в растении – пассивный транспорт
осмос – диффузия растворителя через полупроницаемую мембрану, например, плазматическую мембрану или эндомембраны

Слайд 16эндоцитоз
экзоцитоз
Удивительно, но сейчас есть данные и об
активном транспорте воды
Это только

Слайд 17Некоторые фундаментальные физические основы процессов движения воды в клетке и организме.
Химический
- Растворимые вещества понижают активность молекул воды, находящихся в клетке.
- Кинетическая энергия молекул воды в клетке и в вакуоли ниже, чем в чистой воде снаружи, в частности, в свободном пространстве.
- Энергетический уровень молекул вещества, который характеризуется скоростью их диффузии, получил название химического потенциала.

Слайд 18Химический потенциал воды μw
выражает максимальное количество внутренней энергии молекул воды, которое
μw – химический потенциал чистой воды (принят равным нулю);
аw – активность молекул воды;
RT – множитель необходимый для перевода активность в единицы энергии.

Слайд 19- Активность чистой воды равна единице, а внутри клетки в растворах
- Таким образом, химический потенциал воды в растворах и в клетке меньший, чем в чистой воде.
- Водный потенциал Ψw выражает способность воды в растворе произвести работу в сравнении с работой, которую производит чистая вода в этих же условиях:
где – парциальный молярный объем воды, т. е. объем 1 моля воды (18,0 см3/моль).
Vw

Слайд 20Водный потенциал характеризует способность воды диффундировать, испаряться или поглощаться (и чем
Ψw имеет размерность энергии, поделенной на объем (что совпадает с размерностью давления).
Его величину выражают в атмосферах или барах (1 атм = 1,013 бар = 105 Па).

Слайд 21- Водный потенциал чистой воды равен нулю.
- При увеличении концентрации
- Водный потенциал раствора всегда меньше, чем чистой воды, поэтому молекулы воды в вакуоли обладают меньшей свободной энергией, меньшим потенциалом, чем находящиеся снаружи клетки.
В соответствии со вторым законом термодинамики процессы переноса веществ и энергии самопроизвольно происходят от более высокого уровня химического потенциала к более низкому, т. е. по градиенту потенциала. Так и вода движется в соответствии с этим законом из области высокого потенциала в область низкого, т.е. направление потока воды определяется падением градиента энергии.
Вода не поглощается клеткой, а поступает в клетку за счет разницы водных потенциалов.

Слайд 22Водный потенциал клетки, ткани или органа, целого растения – величина интегральная
Он состоит из осмотического Ψр, матричного Ψw, тургорного Ψt и гравитационного Ψg потенциалов:
Ψw = Ψр+ Ψm+ Ψt + Ψg
. Он состоит из")
Слайд 23Матричный потенциал Ψm
Связан со снижением активности воды за счет гидратации
Определяется слабыми взаимодействиями (нековалетными). Большую роль играют каппилярные силы (образование менисков в почве).
Величина матричного потенциала определяется силами адсорбции между биополимерами цитоплазмы и молекулами воды, т. е. Ψm отражает влияние макромолекул полимеров на активность воды.

Слайд 24Осмотический потенциал Ψр обусловлен наличием в клетках осмотически активных веществ и
Осмотическое давление: Р = iСRT
где С – концентрация раствора в молях;
Т – абсолютная температура;
R – газовая постоянная;
i – изотонический коэффициент, равный 1 + α·(n–1);
– степень ионизации;
n – количество ионов, на которое диссоциирует молекула электролита.

Слайд 25Тургорный потенциал Ψt обусловлен эластичным противодействием клеточной оболочки разбуханию и характеризует
Гравитационный потенциал Ψg отражает влияние на активность воды сил притяжения.
Величины Ψр, Ψm и Ψg всегда отрицательные, так как присутствие растворенных веществ, биополимеров, а также действие сил тяжести снижают активность воды. Ψt, наоборот, положительный, поскольку при действии на воду механического давления (тургорного или гидрастатического) активность молекул воды увеличивается.

Слайд 26Поскольку в большинстве случаев величины как матричного, так и гравитационного потенциалов
Ψw = Ψр+ Ψt
а водный потенциал клетки будет:
Ψкл = –Ψр – Ψt
при Ψр = Ψt клетка не будет
поглощать воду (Р = Т).

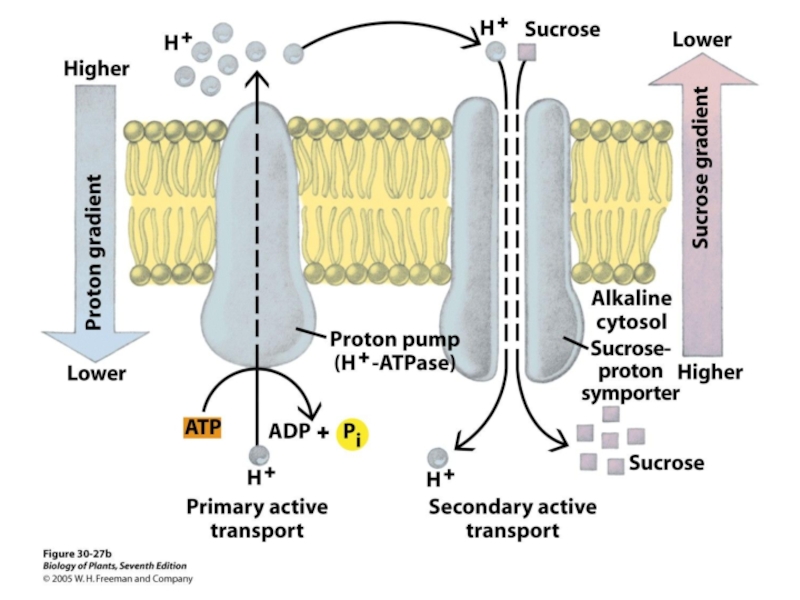
Слайд 27Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины,
формирующие
поэтому другое их название – водные каналы
лат. agua – вода и poros – отверстие


Слайд 29Транспорт воды осуществляют специальные белки аквапорины, формирующие в мембране селективные для
Название аквапорины произошло от лат. agua – вода и poros – отверстие. У растений это 1-2% от общего экстрагируемого белка. Принадлежат к MIP-группе трансмембранных белков (около 150 генов), куда также относятся транспортеры глицерина и др. веществ.
Плазматическая мембрана - семейство plasma mem-
brane intrinsic protein(s) - PIP(s).
Тонопластные водные каналы – отдельное семейство (немного другая структура) – tonoplast intrinsic protein(s) - TIP(s)
Арабидопсис – 7 PIPs + 3 TIPs
Табак – 1 PIPs + 2 TIPs
Бобы – 1 TIP
Подсолнечник – 2 TIPs


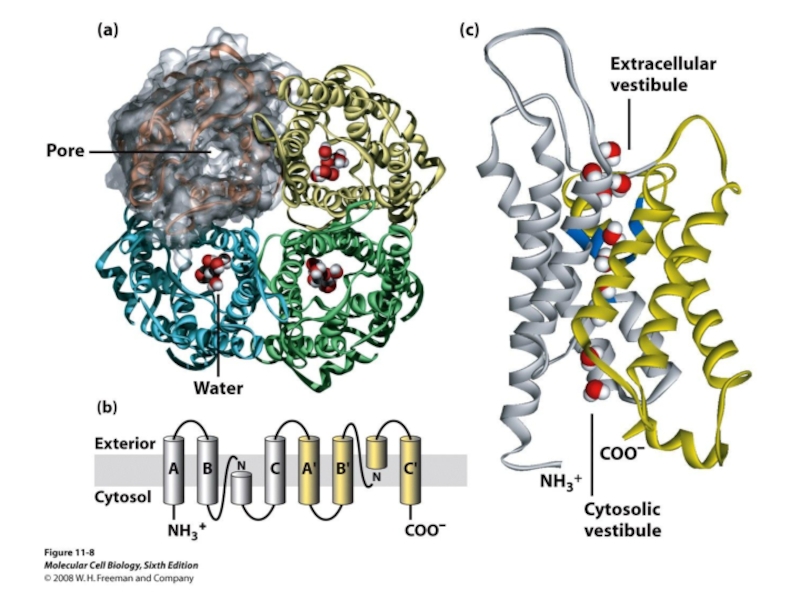
Слайд 31снаружи
цитоплазма
Селективная к воде пора
Одна субъединица
1 нм
Структура аквапорина:
- тетрамерное строение, т.е. состоит
- каждая субъединица имеет 6 трансмембранных доменов (α-спирального строения), расположенных как бы на встречу друг другу, что, как считается способствует транспорту воды в обоих направлениях
Chang et al., 1997, Nature 387:627

Слайд 32
Аквапорины регулируются фосфорилированием как минимум трех сайтов.
Фосфорилирование приводит к активации аквапорина.





Слайд 38
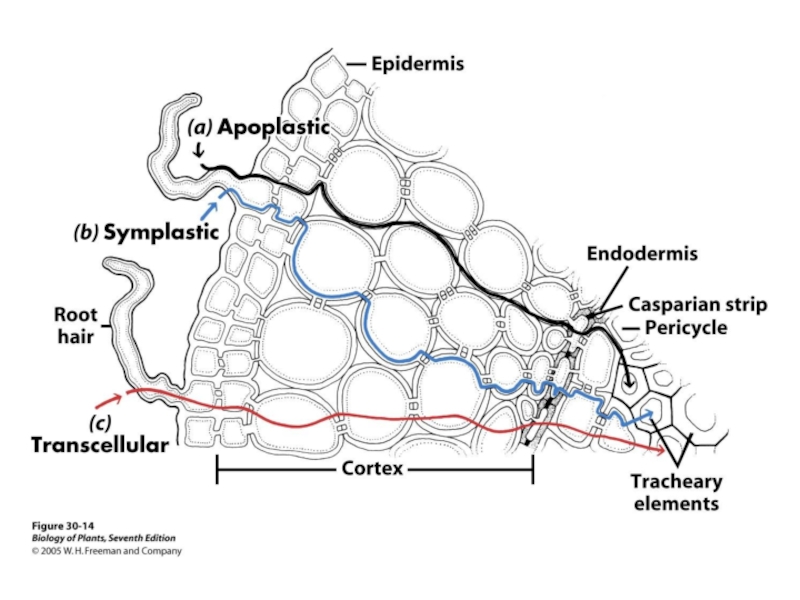
Апопласти-ческий путь
Поясок Каспари
Клетка эндодермы
Симпласти-ческий путь
Поясок Каспари
Плазматическая мембрана
Апопластический путь
Симпласти-ческий путь
Корневой волосок
эпидермис
кортэкс
эндодермис
Сосуды ксилемы
Стела
Вода и растворенные вещества двигаются по симпластическом и апопластическому путям
симпласт – система соединенных живых клеток
апопласт – система соединенных клеточных стенок


Слайд 40Транспорт через плазматическую мембрану эпидермальных клеток корня является началом пути ионов
100 μм
Корень Arabidopsis thaliana

Слайд 41Эпидермальные клетки представлены (примерно 1:1)
трихобластами (образующими корневые волоски) и атрихобластами (необразующими
100 μм
Зрелый
эпидермис
корня
Arabidopsis thaliana
трихобласт
атрихобласт
трихобластами (образующими корневые волоски) и атрихобластами (необразующими корневые волоски)100 μмЗрелый эпидермис")
Слайд 42Листья
(верхний концевой «двигатель»)
Градиент воды
(направление осмоса и корневого давления)
Градиент органических веществ
(направление диффузии)
Почему
Корни
(нижний концевой «двигатель»)
Градиент воды(направление осмоса и корневого давления)Градиент органических веществ(направление диффузии)Почему происходит восходящий ток воды?Корни(нижний")
Слайд 43Гуттация, “слёзы растений”, это выделение воды из листьев
при избыточном корневом давлении

Слайд 44воздух ψ
= −100.0 Mпa
лист ψ (возд. пр.)
=
лист ψ (кл.ст.)
= −1.0 Mпa
Ксилема ствола ψ
= −0.8 Mpa
Ксилема корня ψ
= −0.6 Mпa
Почва ψ
= −0.3 Mпa
Ксилемный сок
Клетки мезофилла
Молекулы воды
транспирация
атмосфера
Адгезия посредством водородных связей
Клеточная стенка
Клетки ксилемы
Когезия и адгезия в ксилеме
Когезия посредством водородных связей
Молекулы воды
Корневой волосок
Почвенная частица
Вода
Поглощение воды из почвы
Градиент водного потенциала
устьица
 = −7.0 Mпaлист ψ (кл.ст.)")
Слайд 45Замыкающие клетки устьиц – главные регуляторы траспирации (присасывающего верхнего концевого двигателя)
Около
Около 95% воды выносится из")
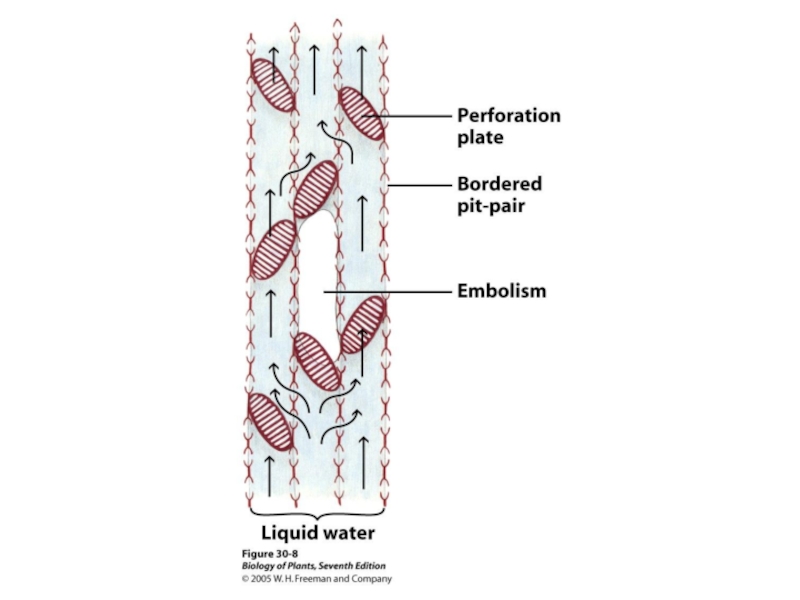
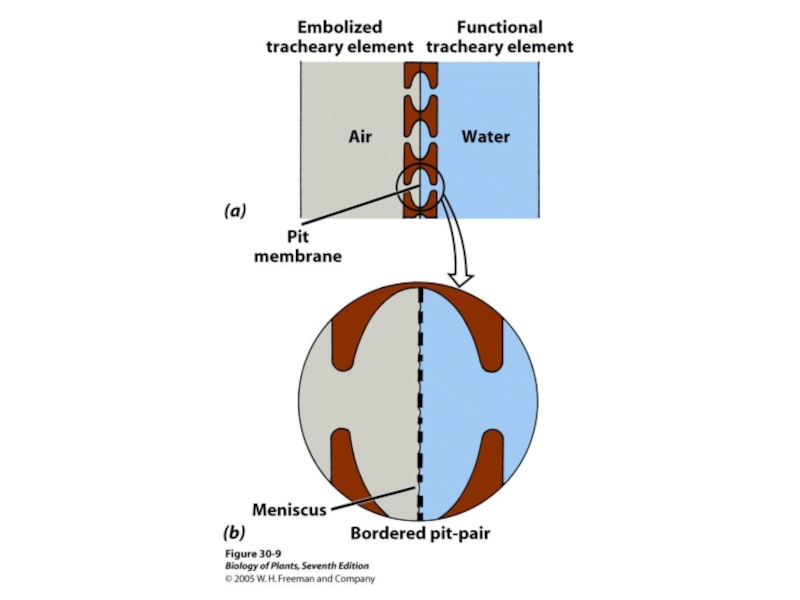
Слайд 46Перфориро-
ванная
пластинка
Сосудистый
элемент
Трахейные (сосудистые элементы
с перфорированными кл. ст. На концах)
трахеиды
отверстия
Трахеиды и
(окраш. С.Э.М.)
сосуд
трахеиды
100 µм
Мертвые клетки!
от греческого ξυλον (ксилон), «дерево"
Ткани ксилемы состоят из трахейных элементов (мертвые клетки), иногда окруженных специализированными живыми паренхимными клетками.
Ксилема - ткань обеспечивающая движение воды
трахеидыотверстияТрахеиды и сосуды(окраш. С.Э.М.)сосудтрахеиды100 µмМертвые клетки!от")
Слайд 47Флоэма
Ситовидно-трубочный
элемент (слева) и
клетка-спутник (Т.Э.М.)
3 мм
Ситовидно-трубочные элементы:
Продольный разрез (С.М.)
Ситовидная
пластинка
Клетки
-спутники
Ситовидно
-трубочный
элемент
плазмодесмы
Ситовидная
пластинка
Ядро клетки
-спутника
Ситовидно-трубочный элемент
Схема продольного разреза
Ситовидная пластинка
с порами (С.Э.М.)
10 µм
30 µм
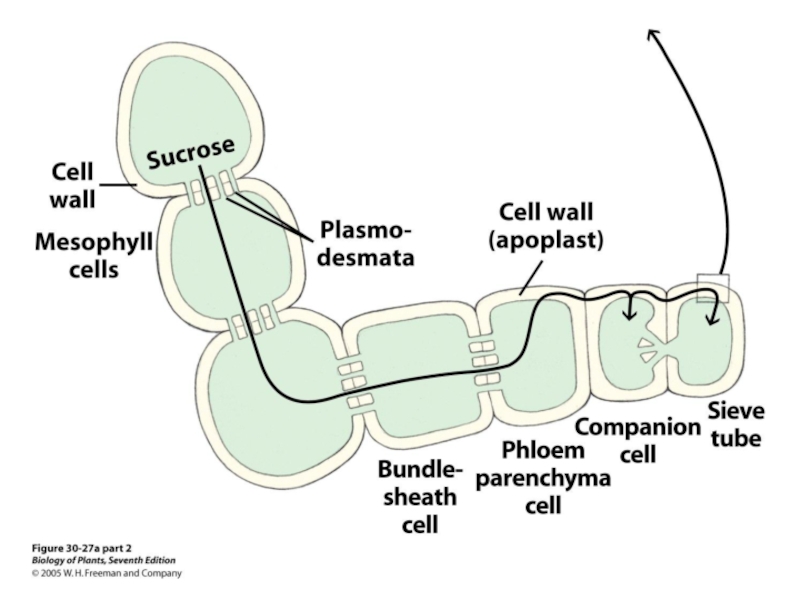
Флоэмная ткань состосит из паренхимных клеток, окруженных ситовидно-трубочными клетками и клетками-спутниками (компаньенами).
от греческого слова φλόος (флоос) «кора".
Живые клетки!
перераспределениеводы по растнию
 и клетка-спутник (Т.Э.М.)3 ммСитовидно-трубочные элементы:Продольный разрез (С.М.)СитовиднаяпластинкаКлетки-спутникиСитовидно-трубочный элементплазмодесмыСитовидная пластинкаЯдро клетки-спутникаСитовидно-трубочный элементСхема продольного")


Слайд 59Транспирация.
Основным органом транспирации является лист. Сверху и снизу лист покрыт
Между нижней и верхней эпидермой находится мезофилл с системой межклетников и проводящими пучками. Межклетники увеличивают внутреннюю испаряющую поверхность листа в 7–10 раз и связываются с окружающей средой через устьица.

Слайд 60Транспирация.
Основным органом транспирации является лист. Сверху и снизу лист покрыт
У большинства растений верхняя эпидерма имеет меньшее количество устьиц (20–100 шт. на 1 мм2 поверхности), чем нижняя (40–400 шт. на 1 мм2 поверхности). Это связано с меньшей тратой воды. Длина устьичной щели – 20–30, а ширина – 4–6 мкм. Обычно устьица занимают 1–2 % площади листа. Однако скорость диффузии водяного пара через устьица довольно велика, поэтому величина испарения с поверхности листа высокая (50–70 % по сравнению с открытым водоемом).


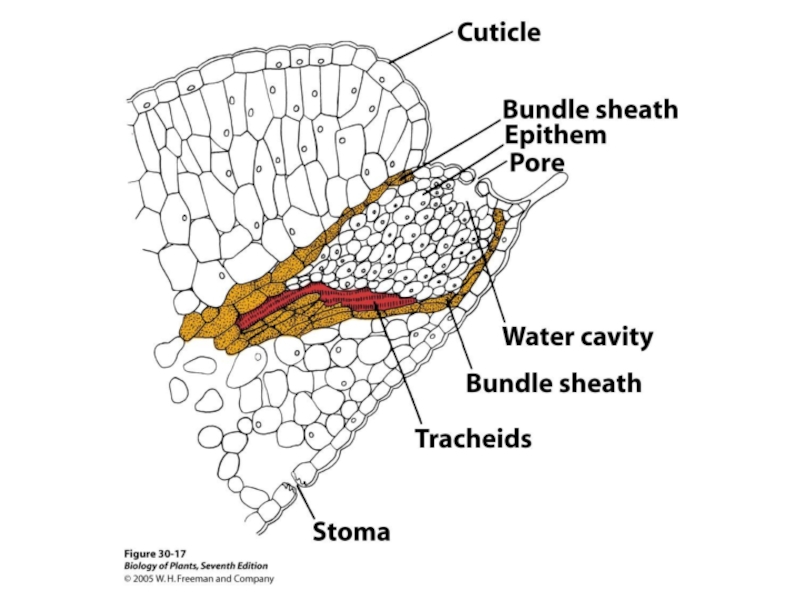
Слайд 62В испарении принимают участие три структуры:
Устьица – поры, через которые
Кутикула – восковой слой, покрывающий эпидермис листьев и стеблей; через нее проходит вода, испаряющаяся с наружных оболочек клеток эпидермиса (около 10 %).
Чечевички, почки – обычно их роль в испарении воды очень мала, но у листопадных деревьев после сбрасывания листьев через них теряется основная масса воды.

Слайд 63Транспирация.
При устьичной транспирации выделяют такие фазы:
– испарение воды с поверхности
– диффузия водяного пара через устьица;
– движение водяного пара с поверхности листа.


Слайд 65Интенсивность (скорость) транспирации определяется количеством граммов воды, испаренной с 1 м2
Обычно скорость транспирации колеблется в интервале от 15 до 250 г/м2·ч, а ночью снижается до 7–20 г/м2·ч. Если провести приблизительные расчеты, то можно показать, что 1 га пашни за счет только транспирации теряет 100 т воды за день.
Продуктивность транспирации показывает число граммов сухого вещества, запасенного в растении при потере 1000 г воды.
Величина продуктивности транспирации варьирует от 1 до 8 г сухого вещества на 1000 г воды.
 транспирации определяется количеством граммов воды, испаренной с 1 м2 листовой поверхности за 1")
Слайд 66Г у т т а ц и я.
Физиологический процесс испарения воды

Слайд 68Системы, которые создают корневое давление и ответственны за поднятие корневого водного
Корневое давление можно измерить, если надетую на перерезанный стебель трубку соединить с манометром. В оптимальных условиях она составляет 2–3 бара.
Количество выделенной пасоки, может отражать поглотительную способность корней.
Начальный восходящий водный ток (корневое давление) обеспечивается живыми клетками, прилегающими к нижнему концу проводящей системы растений – это клетки паренхимы корней – часто эти клетки считают «нижним концевым двигателем».
 вверх по")
Слайд 69Верхний концевой двигатель – системы, обеспечивающие присасывающую силу листьев вследствие испарения
Атмосфера обычно недонасыщена водяными парами, поэтому имеет отрицательный водный потенциал.
При относительной влажности воздуха 90 % он составляет 140 бар.
У большинства растений водный потенциал листьев колеблется от 1 до 30 бар.
По причине большой разности водных потенциалов происходит транспирация.
Уменьшение количества воды в паренхимной клетке листа вызывает снижение активности воды в ней и уменьшение водного потенциала.
Открытые устьица
Закрытые устьица

Слайд 70Воздух
Ψ = −100.0 Mpa
Лист (межклетник)
Ψ = −7.0 Mpa
Ксилемный сок (пасока)
Клетки
Молекулы воды
Транспи-рация
Атмосфера
Адгезия вследствие водородных связей
Клеточная стенка
Клетки ксилемы
Когезия и адгезия в ксилеме
Когезия вследствие водородных связей
Молекулы воды
Корневой волосок
Почвенная частица
Вода
Поглощение воды в корне
градиент водного потенциала
Устьица
Лист (кл. стенка)
Ψ = −1.0 Mpa
Ксилема ствола
Ψ = −0.8 Mpa
Ксилема ствола (подз.)
Ψ = −0.6 Mpa
Почва
Ψ = −0.3 Mpa
Ψ = −7.0 MpaКсилемный сок (пасока)Клетки мезофиллаМолекулы водыТранспи-рацияАтмосфераАдгезия вследствие водородных")
Слайд 711 – вода обладает особой структурой молекулы, которая обеспечивает высокую степень
2 – осмотические явления и изменение водного потенциала лежат в основе транспорта воды по растению
Заключение по лекции
3 – водные каналы представляют собой специализированные белки, участвующие в транспорте воды
4 – симпластический и апопластический пути участвуют в переносе воды в растении по градиенту химического потенциала данного вещества

Слайд 72Самостоятельно – по учебнику
регуляция работы устьичного аппарата экзогенными и эндогенными факторами
.")