- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Физиология мышечной системы презентация
Содержание
- 1. Физиология мышечной системы
- 2. Потенциал действия, его фазы. Соотношение фаз возбудимости с фазами ПД.
- 3. Потенциал действия Потенциал действия — быстрое изменение
- 4. ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ Екр Ео 1 2
- 5. 1) Местное возбуждение (локальный ответ, начальная деполяризация)
- 6. Критический уровень деполяризации (КУД, Екр) тот уровень,
- 7. 2) Деполяризация (восходящая фаза ПД) проницаемость мембраны
- 8. Деполяризация (2) В результате проникновения ионов натрия
- 9. 3)Реполяризация Открыты калиевые каналы, преобладает выходящий
- 10. 4)Следовая деполяризация Отрицательный следовой потенциал Связана с резким снижением калиевого тока и остаточным током натрия
- 11. 5)Следовая гиперполяризация Положительный следовой потенциал Связан с преобладанием выходящего калиевого тока
- 12. СООТНОШЕНИЕ ФАЗ ПД И ВОЗБУДИМОСТИ I -
- 13. Локальный ответ возбудимость ткани повышена( фаза первичной
- 14. фаза деполяризации ПД В период развития фазы
- 15. фаза реполяризации ПД Соответствует фазе относительной рефрактерности
- 16. фаза следовой деполяризации соответствует фазе вторичной экзальтации
- 17. Следовая гиперполяризация Соответствует фазе субнормальной (пониженной)
- 18. Физиология мышц
- 19. Виды мышц • поперечно-полосатые скелетные мышцы;
- 20. Мышечная клетка (мышечное волокно) – вытянутой формы,
- 22. Сократительные белки мышечных клеток – филаменты тонкие
- 23. саркомеры – сократительная
- 26. Тонкие филаменты Закреплены в области Z
- 27. в покое места связывания миозина на
- 29. Миозиновые филламенты образованы повторяющимися молекулами белка
- 32. Пузырьки на концах продольных трубочек - терминальные
- 34. Саркоплазматический ретикулум форма ЭР
- 35. рецептор рианодина в мембране цистерны связан с
- 36. НЕЙРО-МЫШЕЧНЫЙ СИНАПС. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ
- 37. Структура нервно-мышечного соединения концевая пластинка двигательного
- 38. Внутри терминального участка аксона находятся везикул- пузырьки,
- 39. Постсинаптическая мембрана часть мембраны мышечного волокна,
- 42. Процесс передачи импульса в нервно-мышечном синапсе 1.
- 43. 2. Ионы Са++ поступают в цитоплазму синаптического
- 44. 3. Молекулы медиатора диффундируют через синаптическую щель
- 45. 4. Присоединение медиатора к рецептору приводит к
- 47. 6. ПКП(потенциал концевой пластинки) создает ток, который
- 48. Медиатором в нервно-мышечном синапсе является ацетилхолин. После
- 49. Два механизма высвобождения медиатора в синапсе:
- 50. Полное слияние везикулы с плазмолеммой
- 51. Синапсины – семейство фосфопротеинов на
- 52. Три пула пузырьков в пресинаптическом пространстве:
- 53. Процессы, происходящие после генерации ПД на сарколемме
- 55. Теория скользящих нитей Согласно теории скольжения нитей,
- 56. В покое в межфибриллярном пространстве низкая [Ca2+
- 57. После выделения Ca2+ из СР (вследствие активации
- 58. Теория скольжения Хаксли Рабочий цикл поперечных
- 61. В цикле поперечных мостиков АТФ выполняет
- 63. Режимы мышечного сокращения В естественных условиях
- 64. В зависимости от частоты приходящих к
- 65. Одиночные мышечные сокращения Развивается после одного потенциала
- 66. Характеристика одиночного мышечного сокращения Изменение длины
- 67. Тетанус (лат. tetanus, от греч. tétanos — напряжение, оцепенение, судорога) При слишком частой стимуляции мышцы
- 68. Во время тетанического сокращения в мышце многократно
- 70. Зубчатый тетанус очередной импульс совпадает с
- 71. Гладкий тетанус следующий импульс действует на
- 74. Оптимум и пессимум частоты Амплитуда
- 75. Пессимумом частоты называют более высокую частоту раздражения,


Слайд 3Потенциал действия
Потенциал действия — быстрое изменение мембранного потенциала во время возбуждения.
ПД
возникает при действии на мембрану раздражителей пороговой и сверхпороговой силы
Значение ПД:
В нервной ткани - основной способ передачи информации,
В мышечной ткани- пусковой механизм сокращения миоцитов,
В железистой ткани - пусковой механизм секреции
ПД распространяется по мембране
Значение ПД:
В нервной ткани - основной способ передачи информации,
В мышечной ткани- пусковой механизм сокращения миоцитов,
В железистой ткани - пусковой механизм секреции
ПД распространяется по мембране

Слайд 4ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Екр
Ео
1
2
3
4
5
1- местное возбуждение (начальная деполяризация)
2 - деполяризация
3 - реполяризация
4
- следовая деполяризация
5 - следовая гиперполяризация
5 - следовая гиперполяризация
овершут
0
мВ
2 - деполяризация3 - реполяризация4 - следовая деполяризация5 -")
Слайд 51) Местное возбуждение (локальный ответ, начальная деполяризация)
Под влиянием раздражителя пороговой или
сверхпороговой величины проницаемость мембраны клетки для ионов натрия возрастает, т.к. открываются натриевые каналы.
Ионы натрия устремляются внутрь клетки, что приводит к уменьшению величины мембранного потенциала покоя - деполяризация мембраны.
Деполяризация развивается медленно до достижения критического уровня.
Ионы натрия устремляются внутрь клетки, что приводит к уменьшению величины мембранного потенциала покоя - деполяризация мембраны.
Деполяризация развивается медленно до достижения критического уровня.
 Местное возбуждение (локальный ответ, начальная деполяризация)Под влиянием раздражителя пороговой или сверхпороговой величины проницаемость мембраны")
Слайд 6Критический уровень деполяризации (КУД, Екр)
тот уровень, при котором деполяризация мембраны принимает
регенеративный (самоусиливающийся) характер, свидетельствующий о развитии ПД.
Раздражитель, деполяризующий мембрану до КУД, называется пороговым.
Раздражитель, деполяризующий мембрану до КУД, называется пороговым.
тот уровень, при котором деполяризация мембраны принимает регенеративный (самоусиливающийся) характер, свидетельствующий")
Слайд 72) Деполяризация (восходящая фаза ПД)
проницаемость мембраны для ионов натрия увеличивается в
500 раз и превышает проницаемость для ионов калия в 20 раз.
преобладает входящий натриевый ток
преобладает входящий натриевый ток
 Деполяризация (восходящая фаза ПД)проницаемость мембраны для ионов натрия увеличивается в 500 раз и превышает")
Слайд 8Деполяризация (2)
В результате проникновения ионов натрия в цитоплазму и их взаимодействия
с анионами разность потенциалов на мембране исчезает, а затем происходит перезарядка клеточной мембраны (инверсия заряда) - внутренняя поверхность мембраны заряжается положительно по отношению к ее наружной, достигая величины +30-(+50) мВ.
Деполяризация мембраны вызывает инактивацию натриевых каналов и открытие калиевых каналов.
Деполяризация мембраны вызывает инактивацию натриевых каналов и открытие калиевых каналов.
В результате проникновения ионов натрия в цитоплазму и их взаимодействия с анионами разность потенциалов")
Слайд 93)Реполяризация
Открыты калиевые каналы,
преобладает выходящий из клетки калиевый ток
Происходит восстановление поляризации
мембраны (из клетки выходит столько калия, сколько вошло натрия)
Натриевые каналы постепенно выходят из состояния инактивации.
Натриевые каналы постепенно выходят из состояния инактивации.
РеполяризацияОткрыты калиевые каналы, преобладает выходящий из клетки калиевый токПроисходит восстановление поляризации мембраны (из клетки выходит")
Слайд 104)Следовая деполяризация
Отрицательный следовой потенциал
Связана с резким снижением калиевого тока и остаточным
током натрия
Следовая деполяризацияОтрицательный следовой потенциалСвязана с резким снижением калиевого тока и остаточным током натрия")
Слайд 115)Следовая гиперполяризация
Положительный следовой потенциал
Связан с преобладанием выходящего калиевого тока
Следовая гиперполяризацияПоложительный следовой потенциалСвязан с преобладанием выходящего калиевого тока")
Слайд 12СООТНОШЕНИЕ ФАЗ ПД И ВОЗБУДИМОСТИ
I - фаза первичной супервозбудимости (экзальтации)
II -
фаза абсолютной рефрактерности
III - фаза относительной рефрактерности
IV - фаза вторичной супервозбудимости (экзальтации)
V - фаза пониженной возбудимости
III - фаза относительной рефрактерности
IV - фаза вторичной супервозбудимости (экзальтации)
V - фаза пониженной возбудимости
II - фаза абсолютной рефрактерностиIII -")
Слайд 13Локальный ответ
возбудимость ткани повышена( фаза первичной экзальтации).
Во время развития локального
ответа мембранный потенциал покоя приближается к критическому уровню деполяризации и для достижения последнего достаточна сила раздражителя меньшая, чем пороговая (подпороговая).
. Во время развития локального ответа мембранный потенциал покоя")
Слайд 14фаза деполяризации ПД
В период развития фазы деполяризации мембрана утрачивает способность отвечать
возбуждением на раздражители даже сверхпороговой силы.
Наступает полная невозбудимость клетки (фаза абсолютной рефрактерности)
вызвана инактивацией натриевых каналов
Наступает полная невозбудимость клетки (фаза абсолютной рефрактерности)
вызвана инактивацией натриевых каналов

Слайд 15фаза реполяризации ПД
Соответствует фазе относительной рефрактерности
Новый ПД в этой фазе можно
получить при действии сверхпороговых раздражителей
Обусловлена тем, что натриевые каналы выходят из состояния инактивации и возбудимость мембраны постепенно восстанавливается до исходного уровня
Обусловлена тем, что натриевые каналы выходят из состояния инактивации и возбудимость мембраны постепенно восстанавливается до исходного уровня

Слайд 16фаза следовой деполяризации
соответствует фазе вторичной экзальтации
Так как мембранный потенциал в
эту фазу ближе к критическому уровню деполяризации, но сравнению с состоянием покои (исходной поляризацией) , то порог раздражения снижен, т. е. возбудимость повышена.
В эту фазу новый ПД можно вызвать субпороговым раздражителем
В эту фазу новый ПД можно вызвать субпороговым раздражителем

Слайд 17Следовая гиперполяризация
Соответствует фазе субнормальной (пониженной) возбудимости
В эту фазу мембранный потенциал
увеличивается (состояние гиперполяризации мембраны), удаляясь от критического уровня деполяризации, порог раздражения повышается и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины.
 возбудимости В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации")

Слайд 19Виды мышц
• поперечно-полосатые скелетные мышцы;
• поперечно-полосатая сердечная мышца;
• гладкие мышцы внутренних
органов, кожи, сосудов.

Слайд 20Мышечная клетка (мышечное волокно) – вытянутой формы, многоядерная (результат слияния многих
недифференцированных клеток), содержит сократительные белки (миофиламенты)
Миофибриллы – органеллы мышечной клетки с белковыми миофиламентами (тонкие и толстые)
Миофибриллы – органеллы мышечной клетки с белковыми миофиламентами (тонкие и толстые)
 – вытянутой формы, многоядерная (результат слияния многих недифференцированных клеток), содержит сократительные")

Слайд 22Сократительные белки мышечных клеток – филаменты
тонкие
актин: сокращение
небулин: длина актина –
Са++-кальмодулин;
тропонин, тропомиозин: сокращение
толстые
миозин: сокращение,
титин – эластичность мышцы
тропонин, тропомиозин: сокращение
толстые
миозин: сокращение,
титин – эластичность мышцы

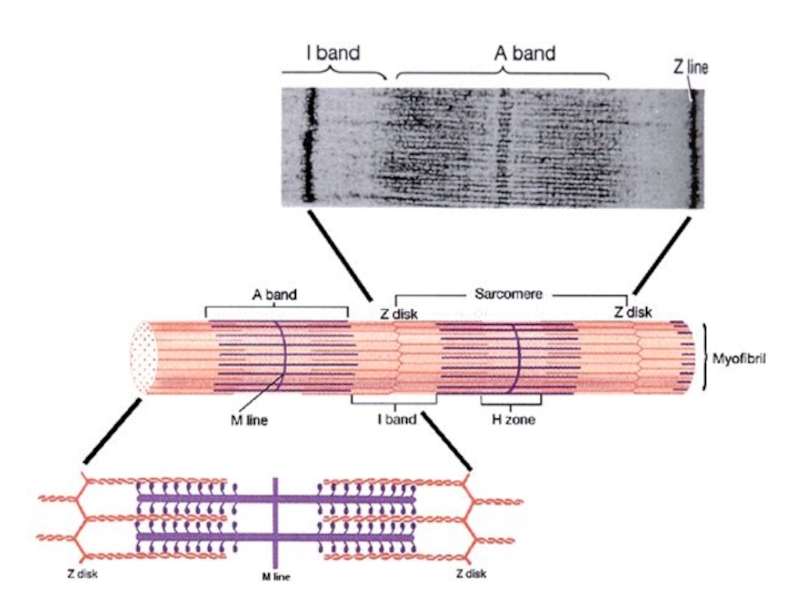
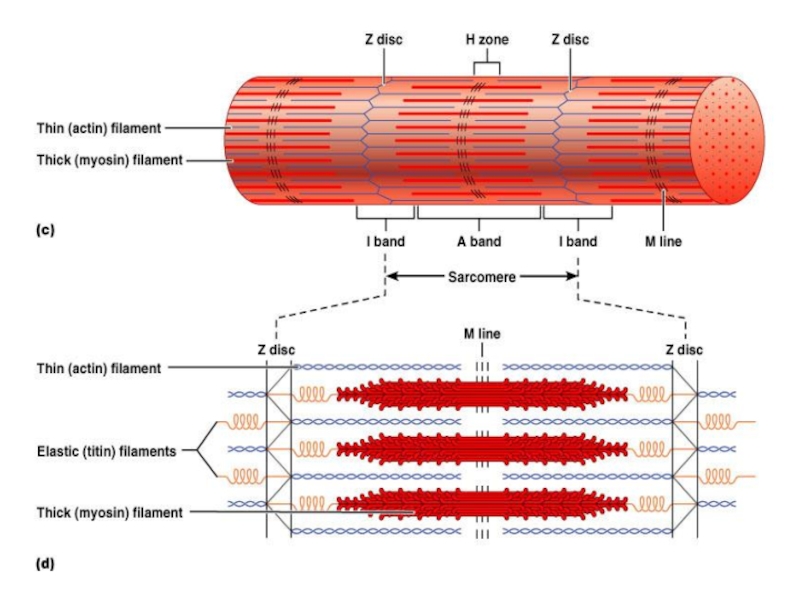
Слайд 23
саркомеры – сократительная единица м. клетки
поперечная исчерченность м.
Z
линии – места связок саркомеров (крепление тонких филаментов)
I – светлые участки – только актин
А – темные участки – миозин +актин
H зона – нет перекрытия актина и миозина
M линия – центр саркомера, где соединяются толстые филаменты
I – светлые участки – только актин
А – темные участки – миозин +актин
H зона – нет перекрытия актина и миозина
M линия – центр саркомера, где соединяются толстые филаменты

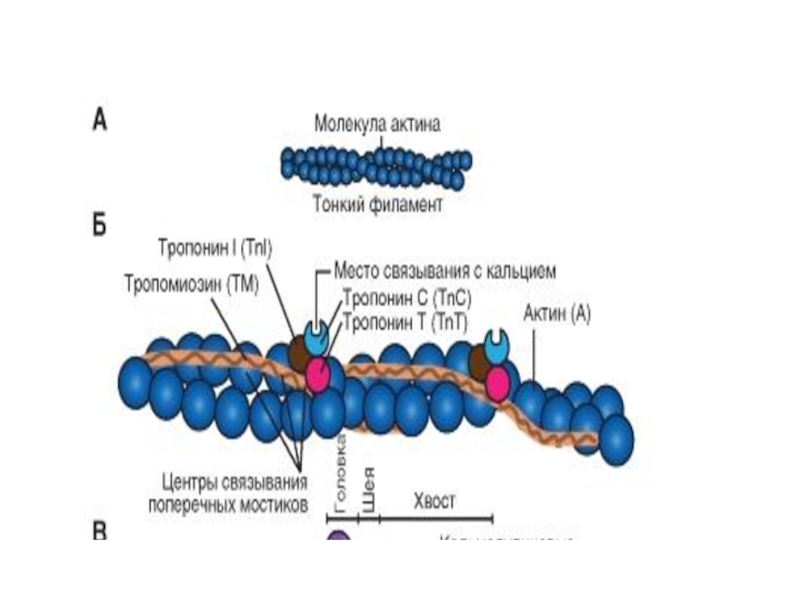
Слайд 26Тонкие филаменты
Закреплены в области Z линии
внешне напоминает две нитки бус,
закрученные в двойную спираль, где каждая бусина – молекула белка актина .
В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.
В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.

Слайд 27
в покое места связывания миозина на актине закрыты тропомиозином,
три формы
тропонина внедрены через регулярные интервалы (тропонин C, I, и T):
тропонин Т связь с тропомиозином,
тропонин I может ингибировать АТФазную активность,
тропонин С обладает значительным сродством к Са2+.
Тропонин в плазме крови – маркер инфаркта миокарда
в норме у здоровых его в плазме нет
при некрозе миокарда – растет уровень в плазме
тропонин Т связь с тропомиозином,
тропонин I может ингибировать АТФазную активность,
тропонин С обладает значительным сродством к Са2+.
Тропонин в плазме крови – маркер инфаркта миокарда
в норме у здоровых его в плазме нет
при некрозе миокарда – растет уровень в плазме

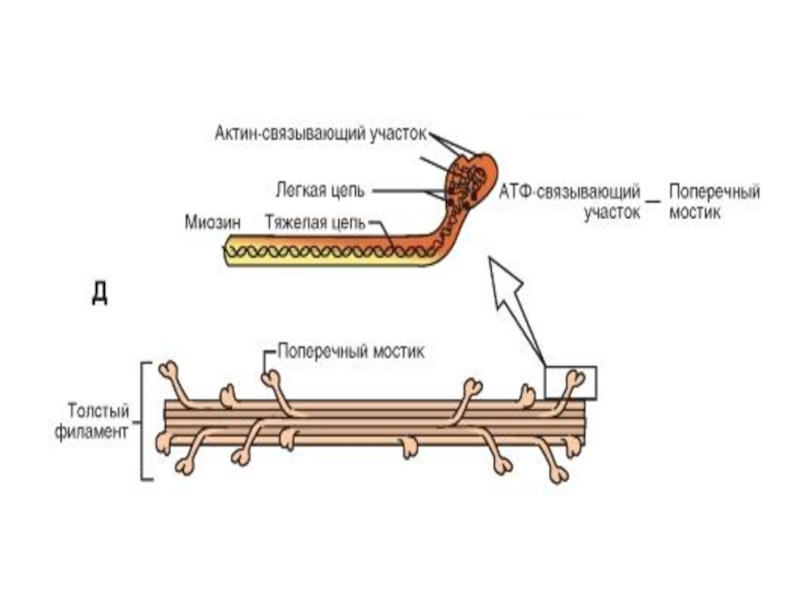
Слайд 29Миозиновые филламенты
образованы повторяющимися молекулами белка миозина .
Каждая молекула миозина
имеет головку и хвост .
Головка миозина имеет 2 участка связывания: -один может связываться с молекулой актина, образуя так называемый поперечный мостик ;
Второй для АТФ (он обладает АТФ-азной активностью.
Головка миозина имеет 2 участка связывания: -один может связываться с молекулой актина, образуя так называемый поперечный мостик ;
Второй для АТФ (он обладает АТФ-азной активностью.

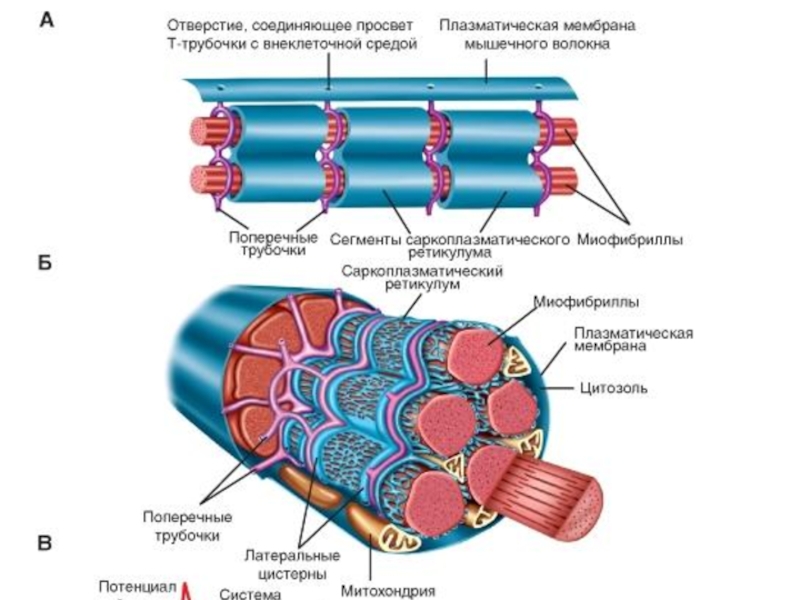
Слайд 32Пузырьки на концах продольных трубочек - терминальные цистерны - подходят очень
близко к поперечным трубочкам, образуя совместно с ними так называемые триады.

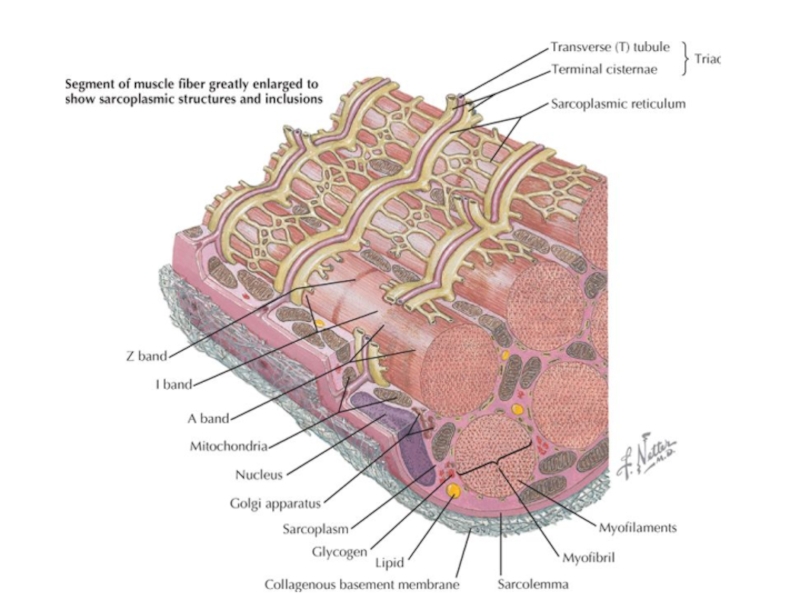
Слайд 34Саркоплазматический ретикулум
форма ЭР
- депо Ca2+
Ca2+ -АТФаза
(закачивает кальций в СР),
Кальсеквестрин (внутри СР) для секвестрации Ca2+,
рианодиновые рецепторы (особый тип Ca2+ каналов) - на мембране СР, взаимодействуют с
- Дигидропиридиновыми рецепторами на плазмолемме
Кальсеквестрин (внутри СР) для секвестрации Ca2+,
рианодиновые рецепторы (особый тип Ca2+ каналов) - на мембране СР, взаимодействуют с
- Дигидропиридиновыми рецепторами на плазмолемме
, Кальсеквестрин (внутри")
Слайд 35рецептор рианодина в мембране цистерны связан с Ca2+-каналом. Работу канала контролируют
рецепторы дигидропиридина плазмолеммы, в ответ на деполяризацию активирующие рецепторы рианодина, что и приводит к высвобождению Ca2+;


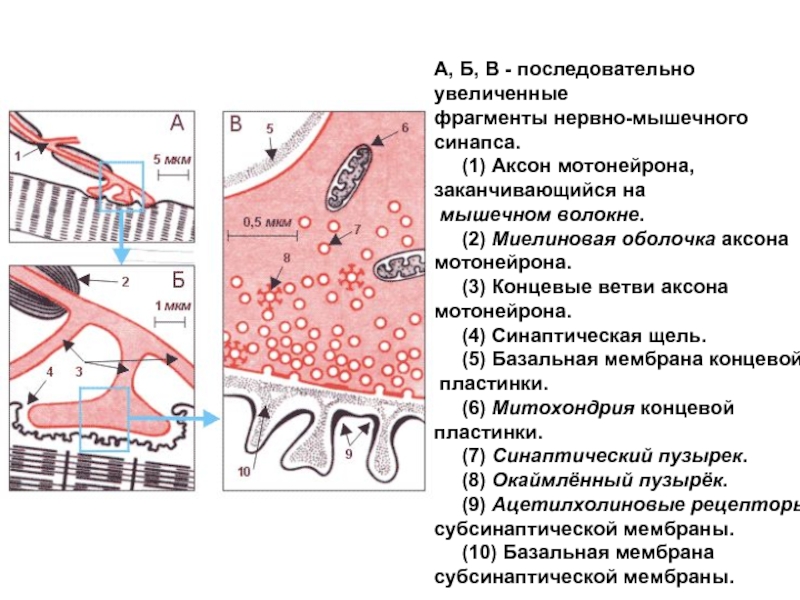
Слайд 37Структура нервно-мышечного соединения
концевая пластинка двигательного нерва на мышце (пресинаптическая мембрана)
концевая мышечная
пластинка (постсинаптическая мембрана)
Синаптическая щель
Синаптическая щель
концевая мышечная пластинка (постсинаптическая мембрана)Синаптическая щель")
Слайд 38Внутри терминального участка аксона находятся везикул- пузырьки, содержащие медиатор- вещество, при
помощи которого передается возбуждение.
Также ферменты, необходимые для синтеза медиатора (синтетазы), митохондрии.
Также ферменты, необходимые для синтеза медиатора (синтетазы), митохондрии.

Слайд 39Постсинаптическая мембрана
часть мембраны мышечного волокна, на которой находятся специальных рецепторы,
чувствительные к определенному медиатору и хемозависимые каналы.

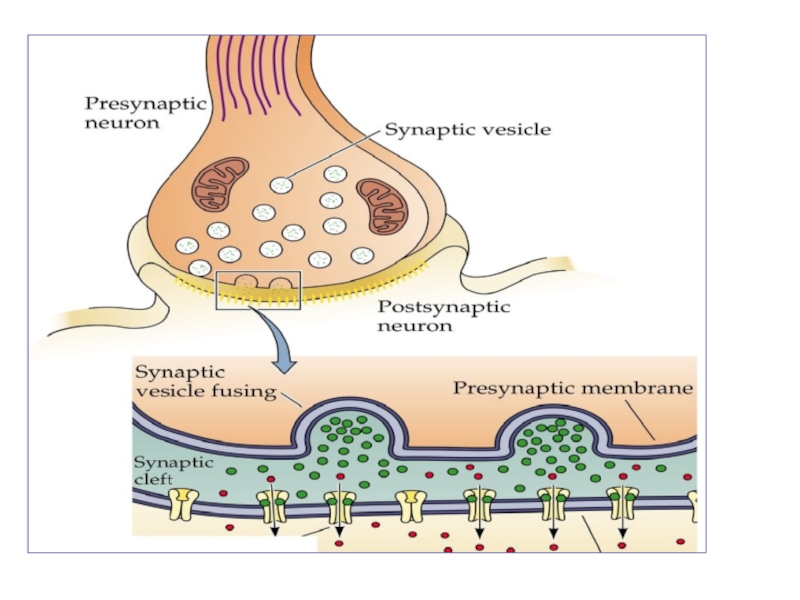
Слайд 42Процесс передачи импульса в нервно-мышечном синапсе
1. Приход в синаптическое окончание возбуждения
приводит к деполяризации пресинаптической мембраны и открытию потенциалчувствительных Са++ -каналов.

Слайд 432. Ионы Са++ поступают в цитоплазму синаптического окончания и активируют опорожнение
синаптических пузырьков в синаптическую щель.
Содержимое синаптических пузырьков поступает в синаптическую щель путем экзоцитоза.
При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10 000 молекул.
Содержимое синаптических пузырьков поступает в синаптическую щель путем экзоцитоза.
При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10 000 молекул.

Слайд 443. Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны,
где связываются с рецепторами хемочувствительных Na+ -каналов.

Слайд 454. Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через
которые в клетку входят ионы Na+.
5. В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП) или потенциал концевой пластинки (ПКП).
5. В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП) или потенциал концевой пластинки (ПКП).

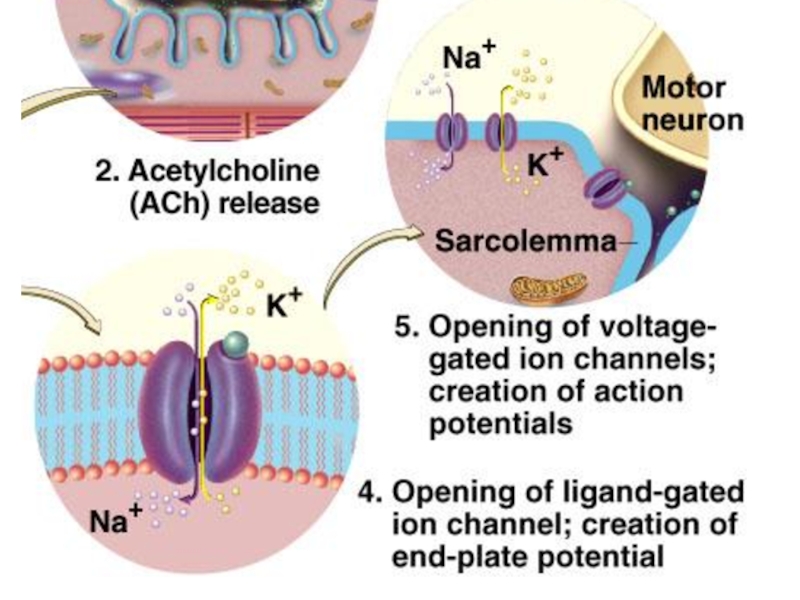
Слайд 476. ПКП(потенциал концевой пластинки) создает ток, который приводит к генерации потенциала
действия на постсинаптической мембране.
 создает ток, который приводит к генерации потенциала действия на постсинаптической мембране.")
Слайд 48Медиатором в нервно-мышечном синапсе является ацетилхолин.
После взаимодействия АХ с рецептором специальный
фермент холинэстераза расщепляет его на составные части, которые могут поступать обратно в пресинаптическое окончание и вновь использоваться для синтеза медиатора.

Слайд 49Два механизма высвобождения медиатора в синапсе:
1.с полным слиянием везикулы с плазмалеммой
и
2.модель «поцеловал и убежал» (kiss-and-run)
везикула соединяется с мембраной,
в синаптическую щель выходит лишь часть содержимого, сама везикула быстро удаляется от мембраны,
связан с ↑↑ Са++ внутри пресинапса
значительно более быстрый процесс
2.модель «поцеловал и убежал» (kiss-and-run)
везикула соединяется с мембраной,
в синаптическую щель выходит лишь часть содержимого, сама везикула быстро удаляется от мембраны,
связан с ↑↑ Са++ внутри пресинапса
значительно более быстрый процесс


Слайд 51Синапсины –
семейство фосфопротеинов на поверхности синаптических пузырьков,
регулируют процесс выброса
нейромедиатора
в дефосфорилированной форме способствуют удержанию пузырьков,
фосфорилирование, является одним из стимулов к выбросу нейромедиаторов.
в дефосфорилированной форме способствуют удержанию пузырьков,
фосфорилирование, является одним из стимулов к выбросу нейромедиаторов.

Слайд 52Три пула пузырьков в пресинаптическом пространстве:
готовый к выбросу,
рециклируемый, и
резервный

Слайд 53Процессы, происходящие после генерации ПД на сарколемме
ПД распространяется по сарколемме в
Т-трубочки
Деполяризация Т-трубочек вызывает открытие потенциал-зависимых Ca2+ каналов и дигидропиридиновых рецепторов, DHPR, которые вызывают возбуждение рианодиновых рецепторов саркоплазматического ретикулума – это вызывает выделение Ca2+ из СР, инициируя сокращение
Деполяризация Т-трубочек вызывает открытие потенциал-зависимых Ca2+ каналов и дигидропиридиновых рецепторов, DHPR, которые вызывают возбуждение рианодиновых рецепторов саркоплазматического ретикулума – это вызывает выделение Ca2+ из СР, инициируя сокращение


Слайд 55Теория скользящих нитей
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему
движению актиновых и миозиновых филламентов друг относительно друга.
Образование поперечных мостиков между актином и миозином происходит при наличии свободного внутриклеточного Ca2+ и доступности АТФ.
Образование поперечных мостиков между актином и миозином происходит при наличии свободного внутриклеточного Ca2+ и доступности АТФ.

Слайд 56В покое в межфибриллярном пространстве низкая [Ca2+ ] 10-7 М - связывание
актина и миозина блокировано тропомиозином
Частично гидролизованная АТФ (АДФ) связана с головками миозина.
Частично гидролизованная АТФ (АДФ) связана с головками миозина.

Слайд 57После выделения Ca2+ из СР (вследствие активации рианодиновых рецепторов) кальций связывается
с тропонином С, устраняется блокирующий эффект тропомиозина, т.к. происходит
открытие на актине участков связывания для миозина
Это создает условия для образования поперечных мостиков
открытие на актине участков связывания для миозина
Это создает условия для образования поперечных мостиков
 кальций связывается с тропонином С, устраняется")
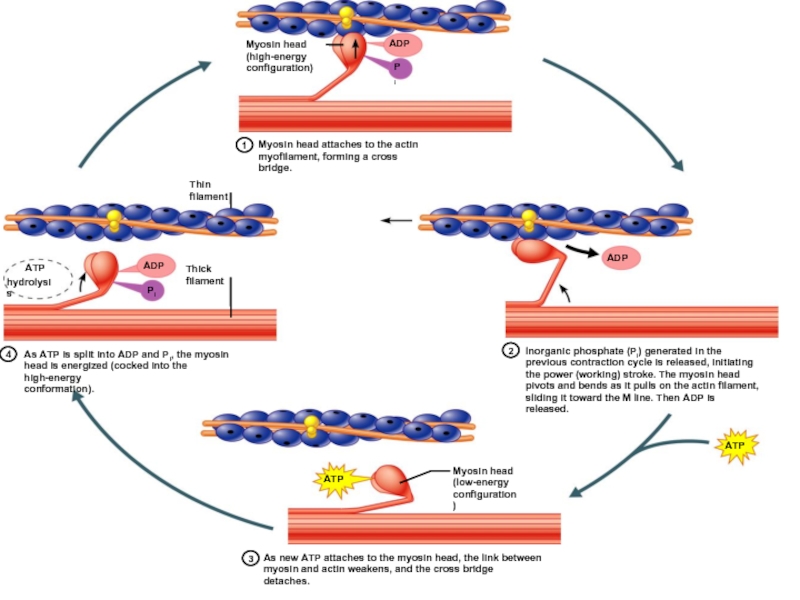
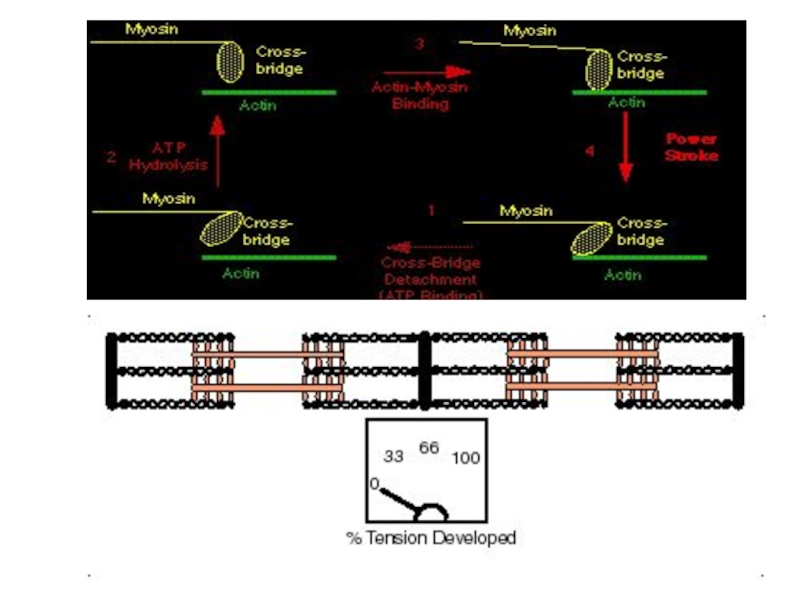
Слайд 58Теория скольжения Хаксли
Рабочий цикл поперечных мостиков:
Образование поперечных мостиков между актином и
миозином (энергия АТФ)
Гребковые движения поперечных мостиков
Скольжение толстых и тонких нитей друг относительно друга
Отсоединение мостиков от актина (затраты энергия АТФ)
перескок мостика на другой участок
Гребковые движения поперечных мостиков
Скольжение толстых и тонких нитей друг относительно друга
Отсоединение мостиков от актина (затраты энергия АТФ)
перескок мостика на другой участок
Гребковые движения")
Слайд 61
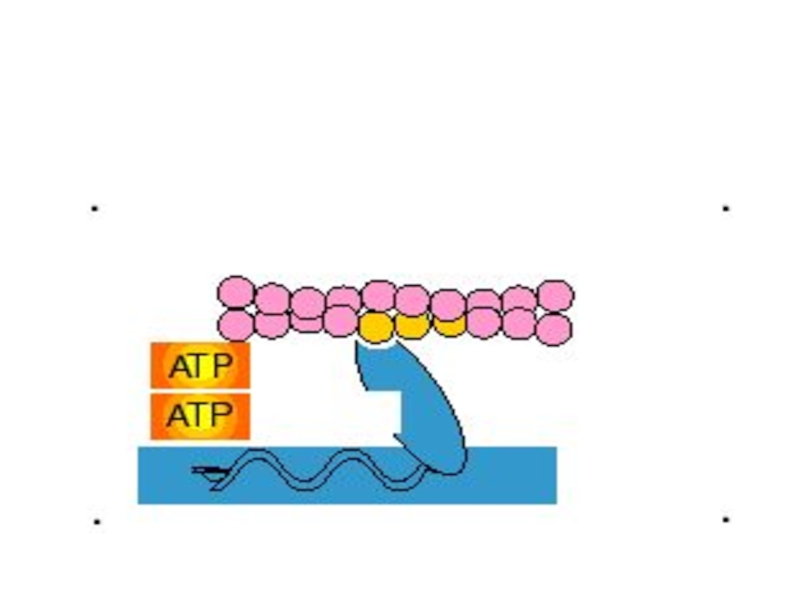
В цикле поперечных мостиков АТФ выполняет две разные роли:
1) гидролиз АТФ поставляет энергию
для движения поперечного мостика;
2) связывание (но не гидролиз) АТФ с миозином сопровождается отделением миозина от актина и создает возможность повторения цикла поперечных мостиков.
2) связывание (но не гидролиз) АТФ с миозином сопровождается отделением миозина от актина и создает возможность повторения цикла поперечных мостиков.
 гидролиз АТФ поставляет энергию для движения поперечного мостика;2) связывание (но")
Слайд 63Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения
не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия.

Слайд 64 В зависимости от частоты приходящих к мышце нервных импульсов мышца может
сокращаться в одном из трех режимов:
• Одиночные мышечные сокращения
зубчатый тетанус
гладкий тетанус

Слайд 65Одиночные мышечные сокращения
Развивается после одного потенциала действия
возникают при низкой частоте электрических
импульсов при стимуляции.
Если очередной импульс (ПД) приходит в мышцу после завершения фазы расслабления.
Если очередной импульс (ПД) приходит в мышцу после завершения фазы расслабления.

Слайд 66Характеристика одиночного мышечного сокращения
Изменение длины мышцы показано синим цветом,
потенциал действия
в мышце - красным,
возбудимость мышцы - фиолетовым.
возбудимость мышцы - фиолетовым.
3 фазы одиночного мышечного сокращения:
• латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
• фаза укорочения (около 50 мс);
• фаза расслабления (около 50 мс).

Слайд 67Тетанус (лат. tetanus, от греч. tétanos — напряжение, оцепенение, судорога)
При слишком частой стимуляции мышцы частотная суммация обусловливает появление
тетануса
- длительного и значительного по амплитуде сокращения мышцы
При слишком частой стимуляции мышцы частотная суммация обусловливает появление тетануса - длительного и значительного")
Слайд 68Во время тетанического сокращения в мышце многократно возникают ПД, обеспечивающие продолжительный
выход ионов Са2+ из саркоплазматического ретикулума и поддерживающие высокий уровень кальция, соединенного с тропонином С.
Цикл образования поперечных мостиков беспрерывно повторяется и сокращение поддерживается до тех пор, пока не прекратится возбуждение.
Цикл образования поперечных мостиков беспрерывно повторяется и сокращение поддерживается до тех пор, пока не прекратится возбуждение.

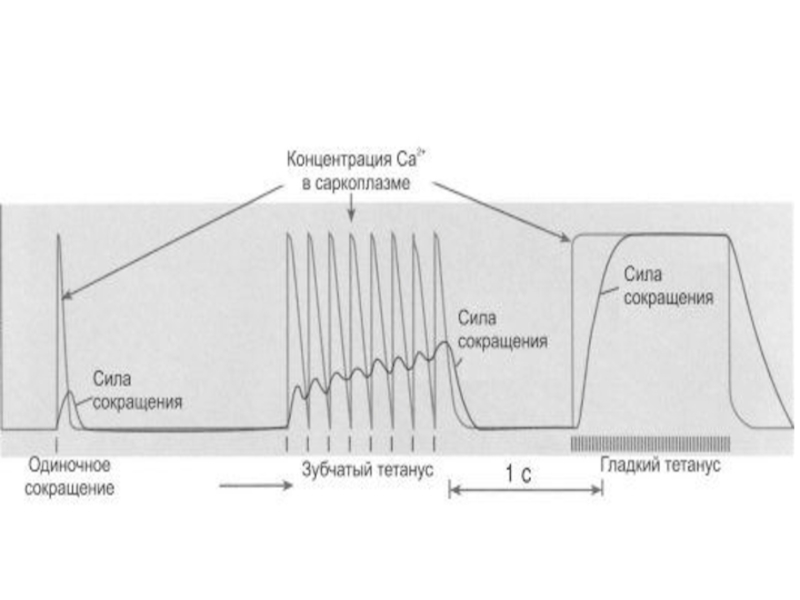
Слайд 70Зубчатый тетанус
очередной импульс совпадает с фазой расслабления предыдущего цикла сокращения.
Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.

Слайд 71Гладкий тетанус
следующий импульс действует на мышцу во время фазы укорочения
(сокращения), в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
, в результате чего")


Слайд 74Оптимум и пессимум частоты
Амплитуда тетанического сокращения, основанного на суммации, зависит
от частоты импульсов, раздражающих мышцу.
Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости и соответственно вызывает тетанус наибольшей амплитуды.
Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости и соответственно вызывает тетанус наибольшей амплитуды.

Слайд 75Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий
импульс тока попадает в фазу рефрактерности, в результате чего амплитуда тетануса значительно уменьшается.