виде конечностей у наземных животных
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Эволюция боковых выростов (отростков), реализующихся в виде конечностей у наземных животных презентация
Содержание
- 1. Эволюция боковых выростов (отростков), реализующихся в виде конечностей у наземных животных
- 2. Эволюционные иновации позвоночных
- 3. Происхождение эволюционных инноваций
- 4. Кости конечности позвоночного. Расположение костей в скелете
- 5. Формирование зачатка конечности начинается с контактов между
- 6. Позиции зачатков конечностей коррелируют с передне-задним паттерном
- 7. Гены гомеотических кластеров Hox-C мыши и HOX-C
- 8. Кластеры гомеотических генов Hox-C мыши и HOX-C
- 9. Экспрессия Hox-генов в мезодерме мышей
- 11. «Гнездовой» способ поддержания активности генов в четырех последовательных участках
- 12. Активность Hox-генов вдоль проксимо-дистальной оси конечности позвоночных
- 13. Последовательное формирование составных частей конечности (стилоподия, зигоподия
- 14. 3 фазы экспрессии Hox-генов в парных конечностях
- 15. Поэтапное формирование конечности от проксимального к дистальному
- 16. Роль генов, содержащих Т-бокс, в развитии конечности
- 17. Распределение транскрипционных факторов Tbx 5 и Tbx4
- 18. Распределение транскрипционных факторов Tbx 5 и Tbx4
- 19. Факторы транскрипции семейства T-бокс требуются для
- 20. Филогенетические связи внутри хордовых, установленные на основании консерватизма T-box-генов
- 22. Происхождение и эволюция конечностей позвоночных
- 24. Как передвигаются позвоночные – от плавников к конечностям
- 25. Эволюция отростков у рыб Акантоды и лопастепёрые
- 26. Двоякодышащие. Слева направо: рогозуб (баррамунда), лепидосирен, большой протоптер (мамба)
- 27. Другой надотряд лопастепёрых – кистепёрые рыбы,
- 28. Латимерия – единственный современный представитель кистепёрых рыб
- 29. Эволюционные модели образования парных выростов тела позвоночных
- 31. Эволюция семейства T-box-генов
- 32. Различия в экспрессии генов Hoxd-11 и Hoxd-13
- 33. Новые данные Показано, что развитие парных отростков
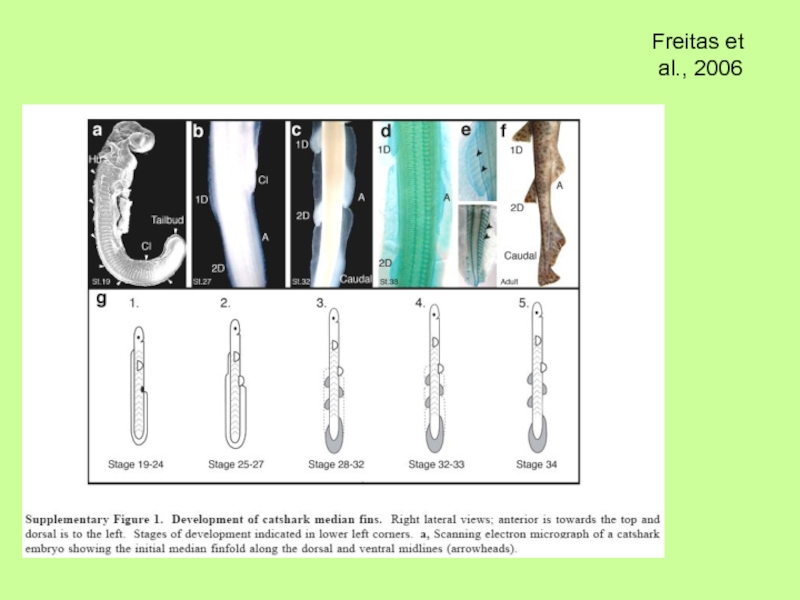
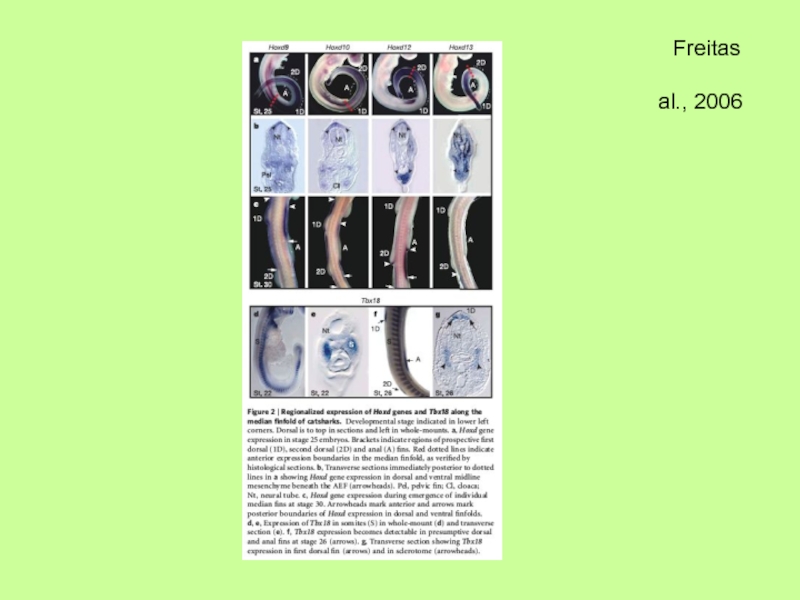
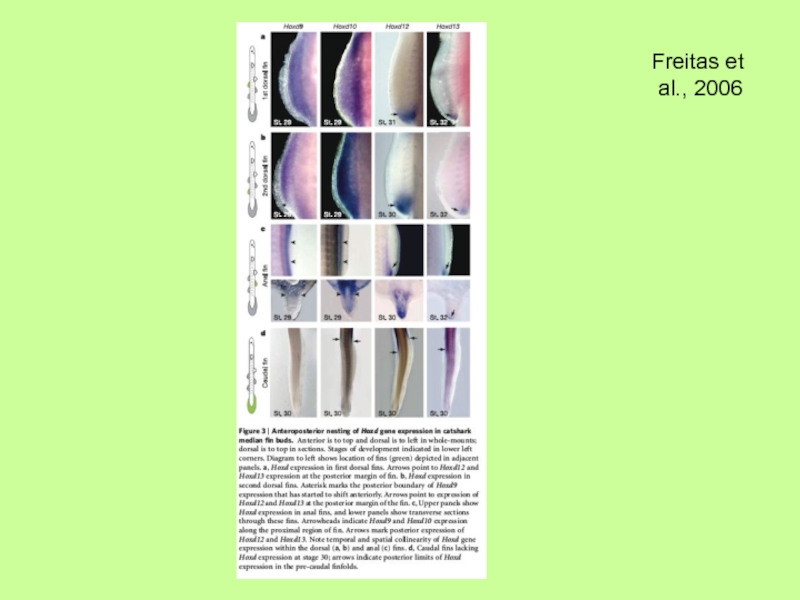
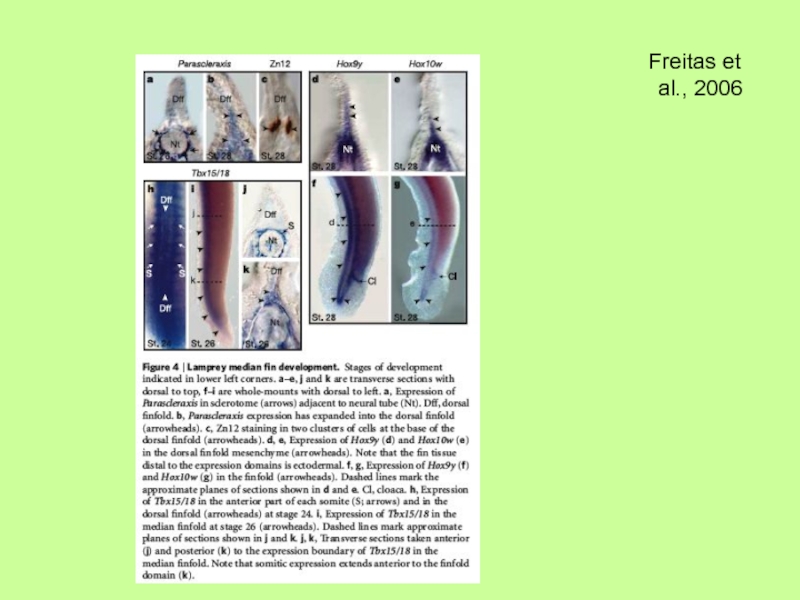
- 36. Молекулярный механизм развития непарных плавников (спинного
- 37. Shubin, Tabin, Carroll, 2009
- 38. АЭГ-стадия в образовании плавника у Danio
Эволюция боковых выростов (отростков), реализующихся в виде конечностей у наземных животных")


Слайд 4Кости конечности позвоночного. Расположение костей в скелете куриного крыла (стилоподий, зигоподий,
аутоподий), направления осей роста конечности (передне-задняя, дорсо-вентральная, проксимо-дистальная)
, направления осей роста")
Слайд 5Формирование зачатка конечности начинается с контактов между эпителиальными клетками эпидермиса, выселяющимися
мезенхимными клетками мезодермы боковой пластинки (предшественниками скелета), мигрирующими клетками митомов (предшественниками скелетной мускулатуры) и клетками нервного гребня (будущая соединительная ткань)

Слайд 6Позиции зачатков конечностей коррелируют с передне-задним паттерном экспрессии Hox-генов. Расположение зачатков
передних конечностей совпадает с передними границами экспрессии Hoxc6 и Hoxb8; Hoxa10 и Hoxd10 – контролируют развитие задних конечностей

Слайд 7Гены гомеотических кластеров Hox-C мыши и HOX-C человека контролируют метамерию заднего
отдела головного мозга и формирование спинного мозга

Слайд 8Кластеры гомеотических генов Hox-C мыши и HOX-C человека контролируют формирование позвоночника.
Их экспрессия определяется градиентом ретиноевой кислоты (возрастающим в передне-заднем направлении).




Слайд 13Последовательное формирование составных частей конечности (стилоподия, зигоподия и аутоподия) млекопитающих определяется
специфическими паттернами экспрессии Hox-генов (4-х паралогичных семейств). Hox-гены в составе кластеров экспрессируются колинеарно формирующимся частям конечности вдоль проксимо-дистальной оси. В дистальной части конечности Hox-гены специфически экспрессируются в передне-заднем направлении (формирование пястья/запястья и пальцев). Мутации Hox-генов ведут к дефектам скелета конечностей.
 млекопитающих определяется специфическими паттернами экспрессии Hox-генов")

Слайд 15Поэтапное формирование конечности от проксимального к дистальному концу (в три фазы)
соответствует последовательной смене специфичных паттернов экспрессии Hox-генов вдоль проксимо-дистальной оси конечности зародыша мыши. В то же время их экспрессия меняется и вдоль передне-задней оси. В первой фазе - экспрессия Hoxd9, Hoxd10 во всей почке . Во второй фазе экспрессия Hox-генов с нумерацией от 9 и больше по гнездовому (матрешечному) типу вдоль передне-задней оси, но Hoxd9 также наибольшая как на заднем, так и и переднем концах . В третьей фазе Hoxd9 инактивируется, но сохраняется передне-заднее нарастание активностей Hoxd10-Hox12 и появляется передне-заднее симметричное распределение активности Hoxa13, Hoxd13-генов.
 соответствует последовательной смене специфичных")

Слайд 17Распределение транскрипционных факторов Tbx 5 и Tbx4 в мезодерме вдоль передне
-задней оси зародыша мыши определяет тип формируемой конечности. Экстра источник FGF10 индуцирует дополнительные конечности (тип которых определяется видом Tbx). Сомиты вдоль оси куриного зародыша визулизированы по экспрессии MRF4 . На перекрывании границ экспрессии Tbx5 и Tbx4- формируется гибридная конечность - крыло-нога

Слайд 18Распределение транскрипционных факторов Tbx 5 и Tbx4 в мезодерме вдоль передне
-задней оси куриного зародыша (соответственно в области передних и задних конечностей) определяет тип конечности. Экстра источник FGF10 индуцирует дополнительные конечности, вид которых определяется типом белков Tbx (Tbx 5 или Tbx4). Tbx5-передняя конечность, Tbx4 -задняя.

Слайд 19
Факторы транскрипции семейства T-бокс требуются для ранних этапов в спецификации клеток,
становящихся на путь дифференцировки, а также для становления общего плана строения тела позвоночного и соответственно органогенеза. T-box (Т-бокс) – район белка, характерен для всех членов этого белкового семейства, необходим и достаточен для специфического связывания упомянутых белков с субпоследовательностью ДНК: 5’-TCACACCT-3’. T-box- довольно протяженный ДНК-связывающий домен, составляет примерно одну третью часть белковой молекулы (17-26 кД).
Гены этих белков были идентифицированы на основании общности, кодируемого ими ДНК-связывающего домена с ДНК-связывающим доменом белкового продукта экспрессии гена Brachyury (T) мыши Mus. musculus. Ген Brachyury был назван по фенотипу его мутации («обрубленный хвост»), идентифицированной 70 лет назад у мышей. Этот ген и его паралоги стали хорошо охарактеризованным семейством, моделью для других животных, которые, как выяснилось, также имели аналогичное семейство генов.
Способный локализоваться в ядре белок Brachyury, содержал характерный ДНК-связывающий домен и проявлял свойства транскрипционного фактора. У гомозиготной мыши выявлялись множественные аномалии развития, приводящие в конце концов к летальному исходу. Для белка Brachyury была постулирована роль фактора транскрипции, регулирующего в дозо-зависимой манере в ходе гаструляции спецификацию и дифференцировку постериорной мезодермы.
T-бокс-содержащие белки мыши специфически появлялись в ходе развития, главным образом, в клетках ряда структур, происходящих из постериорной мезодермы, в ходе образования самой хорды и, затем, у её производных.
Гены этих белков были идентифицированы на основании общности, кодируемого ими ДНК-связывающего домена с ДНК-связывающим доменом белкового продукта экспрессии гена Brachyury (T) мыши Mus. musculus. Ген Brachyury был назван по фенотипу его мутации («обрубленный хвост»), идентифицированной 70 лет назад у мышей. Этот ген и его паралоги стали хорошо охарактеризованным семейством, моделью для других животных, которые, как выяснилось, также имели аналогичное семейство генов.
Способный локализоваться в ядре белок Brachyury, содержал характерный ДНК-связывающий домен и проявлял свойства транскрипционного фактора. У гомозиготной мыши выявлялись множественные аномалии развития, приводящие в конце концов к летальному исходу. Для белка Brachyury была постулирована роль фактора транскрипции, регулирующего в дозо-зависимой манере в ходе гаструляции спецификацию и дифференцировку постериорной мезодермы.
T-бокс-содержащие белки мыши специфически появлялись в ходе развития, главным образом, в клетках ряда структур, происходящих из постериорной мезодермы, в ходе образования самой хорды и, затем, у её производных.

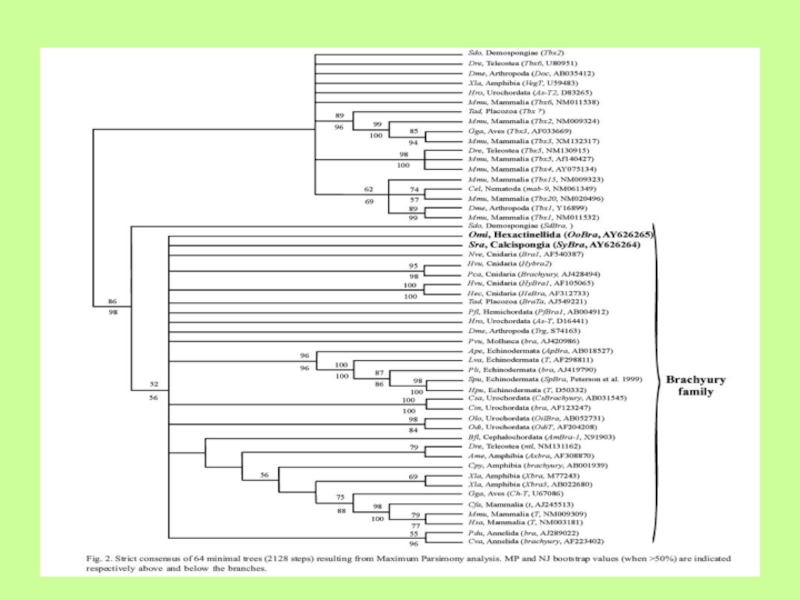
Слайд 20Филогенетические связи внутри хордовых, установленные на основании консерватизма T-box-генов



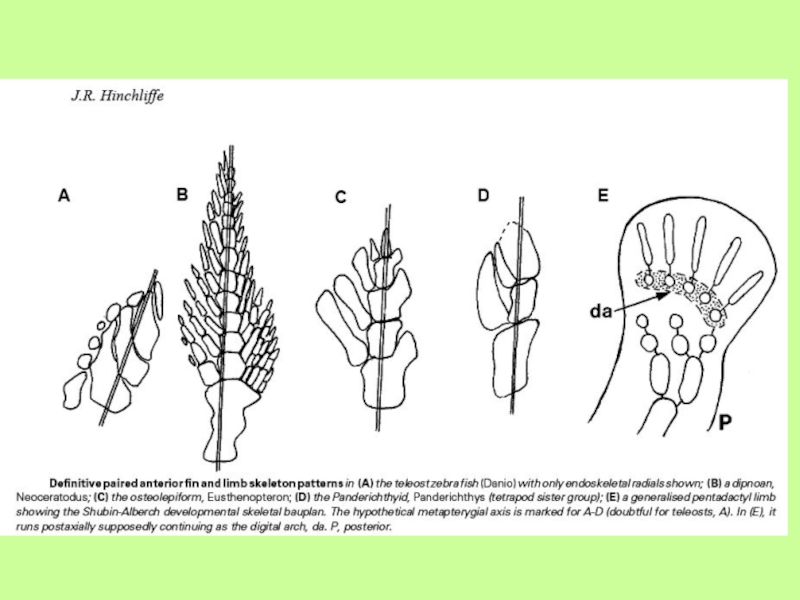

Слайд 25Эволюция отростков у рыб
Акантоды и лопастепёрые рыбы
По-видимому, первыми костными рыбами
были акантоды (Acanthodei), произошедшие в конце силура от панцирных рыб. В настоящее время акантод часто выделяют в отдельный класс. Как правило, это были мелкие веретеновидные рыбы с ганоидной чешуёй. На голове акантод находились мелкие пластинки, перед плавниками – шипы. Акантоды обитали в основном в пресных водах; одни питались планктоном, другие были хищниками.
Акантоды вымерли в начале перми. Но до этого, ещё в девоне от них произошли двоякодышащие рыбы – первые настоящие костные рыбы. Класс костистых рыб делится на два подкласса (иногда выделяемых в отдельные классы): лопастепёрые (Sarcopterygii) и лучепёрые (Actinopterygii). К последним относятся палеониски, хрящевые и костные ганоиды, а также костистые рыбы –Лопастепёрые рыбы подразделяются на кистепёрых и двоякодышащих.
Двоякодышащие рыбы известны прежде всего тем, что помимо жаберного дыхания у них развито и лёгочное, вместо плавательного пузыря имеется своеобразное «лёгкое», соединённое с пищеводом. Некоторые двоякодышащие, поднимаясь на поверхность, могут заглатывать атмосферный воздух. Удлинённое тело может достигать в длину 2 м. Эти рыбы могут пережидать длительную засуху, зарывшись в ил. Непарные плавники имеют характерное для подкласса перистое строение. Верхняя челюсть сращена с черепом, хорда сохраняется всю жизнь. В сердце имеется артериальный конус, между левым и правым предсердиями имеется частичная перегородка. К лёгкому подходит лёгочная дуга кровообращения. Кишечник со спиральным клапаном открывается в клоаку.
Двоякодышащие – пресноводные рыбы, обитающие в стоячих или пересыхающих водоёмах. К настоящему времени сохранилось только 3 семейства этих животных отряда рогозубообразных (6 видов): рогозуб в Австралии, лепидосирен в бассейне Амазонки и протоптеры в Центральной Африке. Все двоякодышащие имеют некоторое промысловое значение.
Акантоды вымерли в начале перми. Но до этого, ещё в девоне от них произошли двоякодышащие рыбы – первые настоящие костные рыбы. Класс костистых рыб делится на два подкласса (иногда выделяемых в отдельные классы): лопастепёрые (Sarcopterygii) и лучепёрые (Actinopterygii). К последним относятся палеониски, хрящевые и костные ганоиды, а также костистые рыбы –Лопастепёрые рыбы подразделяются на кистепёрых и двоякодышащих.
Двоякодышащие рыбы известны прежде всего тем, что помимо жаберного дыхания у них развито и лёгочное, вместо плавательного пузыря имеется своеобразное «лёгкое», соединённое с пищеводом. Некоторые двоякодышащие, поднимаясь на поверхность, могут заглатывать атмосферный воздух. Удлинённое тело может достигать в длину 2 м. Эти рыбы могут пережидать длительную засуху, зарывшись в ил. Непарные плавники имеют характерное для подкласса перистое строение. Верхняя челюсть сращена с черепом, хорда сохраняется всю жизнь. В сердце имеется артериальный конус, между левым и правым предсердиями имеется частичная перегородка. К лёгкому подходит лёгочная дуга кровообращения. Кишечник со спиральным клапаном открывается в клоаку.
Двоякодышащие – пресноводные рыбы, обитающие в стоячих или пересыхающих водоёмах. К настоящему времени сохранилось только 3 семейства этих животных отряда рогозубообразных (6 видов): рогозуб в Австралии, лепидосирен в бассейне Амазонки и протоптеры в Центральной Африке. Все двоякодышащие имеют некоторое промысловое значение.
, произошедшие")
, лепидосирен, большой протоптер (мамба)")
Слайд 27
Другой надотряд лопастепёрых – кистепёрые рыбы, произошедшие в девоне от древних
двоякодышащих. Они ползают по дну, опираясь на мускулистые парные плавники, укреплённые кистеобразно разветвляющимися скелетными фрагментами подобно конечностям наземных позвоночных. Спинных плавников два. Череп разделён на две части, подвижные друг относительно друга. Хорда сохраняется всю жизнь.
Все кистепёрые рыбы – хищники. Вымершие в начале перми рипидистиобразные имели внутренние ноздри, что позволило им выбраться на сушу и стать родоначальниками земноводных. Древние целакантообразные до недавнего времени также считались вымершими, поэтому открытие в 1938 году у Коморских островов живого целаканта – латимерии – стало настоящей сенсацией, которую можно сравнить разве что с поимкой живого динозавра. Латимерии – крупные рыбы длиной более 1,5 м и массой до 100 кг. Эти животные никогда не выходили на сушу и потому снова утратили внутренние ноздри и лёгкие.
Все кистепёрые рыбы – хищники. Вымершие в начале перми рипидистиобразные имели внутренние ноздри, что позволило им выбраться на сушу и стать родоначальниками земноводных. Древние целакантообразные до недавнего времени также считались вымершими, поэтому открытие в 1938 году у Коморских островов живого целаканта – латимерии – стало настоящей сенсацией, которую можно сравнить разве что с поимкой живого динозавра. Латимерии – крупные рыбы длиной более 1,5 м и массой до 100 кг. Эти животные никогда не выходили на сушу и потому снова утратили внутренние ноздри и лёгкие.




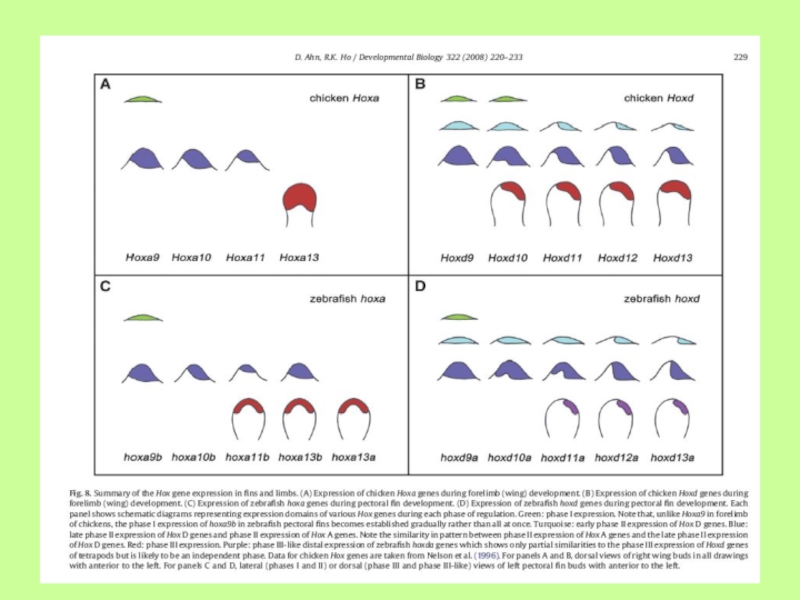
Слайд 32Различия в экспрессии генов Hoxd-11 и Hoxd-13 в зачатке плавника рыб
и конечности четвероного. Описанного ранее паттерна экспрессии Hox-генов в конечностях, по-видимому, нет в плавнике рыбы (аналоге конечности), что свидетельствует об эволюционных изменениях в паттерне экспрессии Hox-генов наземных позвоночных (по данным 90-х годов ХХ века)

Слайд 33Новые данные
Показано, что развитие парных отростков костистых рыб происходит в манере
3-х фазной (бифазной) активации Hox-генов в проксимо-дистальном и в передне-заднем направлениях и в этом смысле мало чем отличается от закладок конечностей наземных животных
 активации")
Слайд 36
Молекулярный механизм развития непарных плавников (спинного и брюшного) свидетельствует, что они
в отличие от парных закладываются из клеток склеротома (параксиальной мезодермы) и в присутствии АЭГ, но также как и парные отростки (скелет из мезодермы боковых пластинок) позвоночных связаны с использованием одних и тех же (сходных?) генетических программ, касающихся действия Hox-генов, Tbx- генов, Shh-гена и т.п.
 свидетельствует, что они в отличие от парных")