- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Этилен презентация
Содержание
- 1. Этилен
- 2. Ethylene concentration µL L-1 (ppm) -pea seedlings
- 3. Выявление «этиленовых» мутантов (по тройному ответу)
- 4. Биосинтез этилена
- 5. Тканеспецифичность экспрессии генов ACS A, etiolated seedlings
- 6. Регуляция
- 7. Фенотипы мутантов eto (ethylene overproducing) и cin
- 8. Уровень этилена в тканях регулируется через стабильность
- 9. Регуляция биосинтеза этилена через стабильность ACS белков ETO1 – субъединица убиквитин-лигазного комплекса CUL3-BTB
- 10. ethylene air
- 11. Передача сигнала при ответе на этилен Основные
- 12. Белки двухкомпонентных систем передачи сигнала у Arabidopsis
- 13. Пять этиленовых рецепторов Arabidopsis Домены: Трансмембранный (ТМ)
- 14. Сравнение систем передачи сигнала цитокинина и этилена
- 15. Гибридные гистидин-киназы CRE1 и CKI (рецепторы цитокинина),
- 16. Мутации с потерей функции одного из этиленовых
- 17. В воздухе (без этилена): Рецепторы связывают и
- 18. Кофакторы этиленовых рецепторов Ионы Cu+ - связывание
- 19. Важные мотивы лиганд-вязывающих ТМ доменов: 1. мотивы
- 20. Ser/Thr протеинкиназа CTR1 (МАРККК) – негативный регулятор
- 21. CTR1 образует олигомеры и вызывает олигомеризацию этиленовых рецепторов J. Mol. Biol.: October 2012 CTR1 рецепторы
- 23. Предполагаемая схема действия МАР-киназного каскада в этиленовом
- 24. Белок EIN2 - центральный регулятор этиленового сигналинга
- 25. Взаимодействие EIN2 с CTR1 и рецепторами этилена
- 26. Этилен-зависимые транскрипционные факторы EIN3 и EIL1
- 27. ТФ EIN3 – координаторы ответа на этилен и его взаимодействия с другими гормонами
- 28. Регуляция стабильности белков EIN3 Стабилизаторы EIN3: MAPK
- 29. Регуляция стабильности белков EIN3 Белки EIN3 подвергаются
- 30. EIN3 EBF genes, etc. EBF
- 31. Рибонуклеаза EIN5/XRN4 регулирует уровень транскриптов EBF1/2 Продукт
- 32. Факты: Сверхэкспрессия 3’UTR генов EBF вызывает нечувствительность
- 33. EIN2 репрессирует трансляцию EBF транскриптов полиU-шпильки
- 34. Полиуридиновые мотивы в 3‘UTR EBF генов
- 35. Опыты по ко-локализации 3‘UTR EBF с белками EIN2 и EIN5
- 36. Транскрипционные факторы ERF Транскрипционные факторы EIN3 и
- 37. Филогенетическое древо ERF белков разных видов растений
- 38. ERF-активаторы и ERF-репрессоры Содержат мотивы EDLL (Глн
- 39. ERFs Абиотический стресс (поранение, высокая температура, высыхание)
- 40. Некоторые ERF - центральные регуляторы защиты от
- 41. Роль этилена в реакции сверхчувствительности (HR)
- 42. Фитогормоны и системная устойчивость растений
- 43. Газообразные гормоны – этилен и метил-жасмонат могут
- 44. Местный (HR) и системный (SAR)
- 45. PAMP-triggered (PTI) and Effector-trigegred (ETI) plant immunity: 2 барьера защиты
- 46. PAMP-triggered (PTI) and Effector-trigegred (ETI) plant immunity: 2 барьера защиты “Gene for gene”
- 47. ТФ NPR1 (nonexpresser of PR genes 1)
- 48. Этилен и созревание плодов
- 49. Климактерические и неклимактерические плоды НЕклимактерические – пик
- 50. Роль этилена и АБК в созревании плодов
- 51. В процессе созревания климактерических плодов повышается биосинтез
- 52. Роль этилена в старении листьев и созревании
- 53. Внешние факторы не вызывают старения Внешние факторы
- 54. Взаимодействие гормонов и ТФ в регуляции возрастозависимого
- 55. Взаимодействие гормонов и ТФ в регуляции старения
- 56. SAG (senescence-associated genes): ТФ семейств NAC, WRKY,
- 57. Гомологи ORE1 есть у всех наземных растений,

Слайд 2Ethylene concentration µL L-1 (ppm)
-pea seedlings
-grown in darkness for 48h
-in the
0
0.012
1.6
0.8
0.06
0.1
0.2
0.025
0.4
Original
size
saturation
Тройной ответ проростков на этилен
Д.Н. Нелюбов
-pea seedlings-grown in darkness for 48h-in the presence of ethylene00.0121.60.80.060.10.20.0250.4OriginalsizesaturationТройной ответ")
Слайд 3
Выявление «этиленовых» мутантов (по тройному ответу)
воздух
этилен
Дикий тип
этилен
Этилен-нечувствительный мутант (ein, etr)
Мутант с
сtr1
этилен
воздухэтиленДикий типэтиленЭтилен-нечувствительный мутант (ein, etr)Мутант с конститутивным тройным ответом (даже")

Слайд 5Тканеспецифичность экспрессии генов ACS
A, etiolated seedlings
B, light grown seedlings
1, roots
2,
3, stems
4, flowers
5, siliques
6, genomic DNA.


Слайд 7Фенотипы мутантов eto (ethylene overproducing) и cin (cytokinin insensitive)
Мутации cin
Мутации eto – результат стабилизации ферментов ACS за счет изменения последовательности их С-концевого домена (eto 2,3) или снижения активности убиквитин-лигазного комплекса (eto 1)
 и cin (cytokinin insensitive) Мутации cin – результат потери функции")
Слайд 8Уровень этилена в тканях регулируется через стабильность ACS белков
А – Деградация
В – Стабилизация ACS белков путем фосфорилирования по консервативным остаткам серина: с помощью Са2+-зависимых киназ CDPK (все ACS) и MAPКК киназы МРК6 (ACS2 и 6).

Слайд 9Регуляция биосинтеза этилена через стабильность ACS белков
ETO1 – субъединица убиквитин-лигазного

Слайд 10ethylene
air
1. Мутанты нечувствительные к этилену (etr1,
2. Мутанты с конститутивным тройным ответом при отсутствии этилена (crt1 – рецессивный).
Компоненты этиленового сигналинга были выявлены при изучении мутантов A.thaliana с измененным «тройным ответом»:
Изучение эпистатического взаимодействия мутаций ? порядок действия генов:
EBF1, 2
ETP
;")
Слайд 11Передача сигнала при ответе на этилен
Основные компоненты:
Рецепторы этилена: ETR1 и 2,
Металлотранспортер RAN
3. Киназа МАРККК CTR1, нижележащие компоненты МАР-киназного каскада
4. Мембранный белок EIN2
5. Транскрипционные факторы EIN3-like
6. Компоненты убиквитин-лигазных комплексов ETP1, 2; EBF1, 2 (действуют на разных этапах сигналинга)
7. 5’-3’-экзорибонуклеаза EIN5
8. Транскрипционные факторы ERF

Слайд 12Белки двухкомпонентных систем передачи сигнала у Arabidopsis
A - Гибридные сенсорные гистидин-киназы
B - ARR– регуляторы ответа (передача сигнала цитокинина): В-типа (ARR2) – транскрипционные факторы и А-типа (ARR4) –репрессоры транскрипции
С - AHP – гистидин-содержащие фосфотрансферазы (передача сигнала цитокинина)
Консервативные а/к остатки: H – His, D - Asp.
 и")
Слайд 13Пять этиленовых рецепторов Arabidopsis
Домены:
Трансмембранный (ТМ) домен – связывает этилен в присутствии
три раза пронизывает мембрану. У рецепторов подсемейства 2 содержит экстраклеточную сигнальную последовательность (не важна для функции)
GAF домен – содержит мотив coiled coil, служит для образования гомо- и гетеродимеров рецепторов
Гистидин-киназный (НК) домен – 1. автофосфорилируется при связывании этилена (киназная активность не важна для функции рецепторов). 2. взаимодействует с CTR1 (важно для функции рецепторов!)
Receiver домен – у ETR1, 2 и EIN4. Не важен для функции рецепторов!
 домен – связывает этилен в присутствии кофактора (Cu 1+)три раза")
Слайд 14Сравнение систем передачи сигнала цитокинина и этилена
Фосфореле
Активация каждого последующего компонента предыдущим
Нет
Инактивация рецепторов при связывании лиганда (этилена)
Плазма-лемма
Мембраны ЭР и АГ

Слайд 15Гибридные гистидин-киназы CRE1 и CKI (рецепторы цитокинина), а также ETR1 (рецептор
Взаимодействие белков, входящих в двухкомпонентные системы передачи сигналов у Arabidopsis:
, а также ETR1 (рецептор этилена) могут передавать фосфатную")
Слайд 16Мутации с потерей функции одного из этиленовых рецепторов не имеют проявления,
Функции рецепторов перекрываются
Рецепторы негативно регулируют ответ на этилен
Рецепторы являются негативными регуляторами этиленового сигналинга
1

Слайд 17В воздухе (без этилена):
Рецепторы связывают и активируют протеинкиназу CTR1, которая негативно
При связывании этилена:
Происходит высвобождение CTR1 ? нет подавления ответа на этилен. Убиквитинирование и протеасомная деградация рецепторов
Инактивация рецепторов при связывании этилена
:Рецепторы связывают и активируют протеинкиназу CTR1, которая негативно регулирует ответ на этиленПри")
Слайд 18Кофакторы этиленовых рецепторов
Ионы Cu+ - связывание этилена происходит только при их
Трансмембранный белок RAN1 (Response to ANtagonist 1) – осуществляет транспорт ионов Cu+. Локализован в аппарате Гольджи. RAN1 действует в сигналинге этилена ВЫШЕ рецепторов
Трансмембранные белки RTE (Reversion to Ethylene senstitivity) арабидопсиса и GR (Green-Ripe) томата – взаимодействуют с рецептором ETR1. Локализованы в аппарате Гольджи и ЭР.

Слайд 19Важные мотивы лиганд-вязывающих ТМ доменов: 1. мотивы из 7 консервативных аминокислот,
«Включение» этиленового сигналинга – массовый переход рецепторов в «состояние 3»
3 трансмембранные α-спирали лиганд-связывающих доменов – важны для связывания иона меди и конформационных изменений

Слайд 20Ser/Thr протеинкиназа CTR1 (МАРККК) – негативный регулятор этиленового сигналинга
ETR
CTR1
Ответ
на этилен
Мишени
MAP-киназный каскад
EIN2 – мембранный белок
EIN3 – этилен-зависимый транскрипционный фактор
 – негативный регулятор этиленового сигналингаETRCTR1Ответ на этиленМишени CTR1:MAP-киназный каскадEIN2 – мембранный")
Слайд 21CTR1 образует олигомеры и вызывает олигомеризацию этиленовых рецепторов
J. Mol. Biol.: October
CTR1
рецепторы

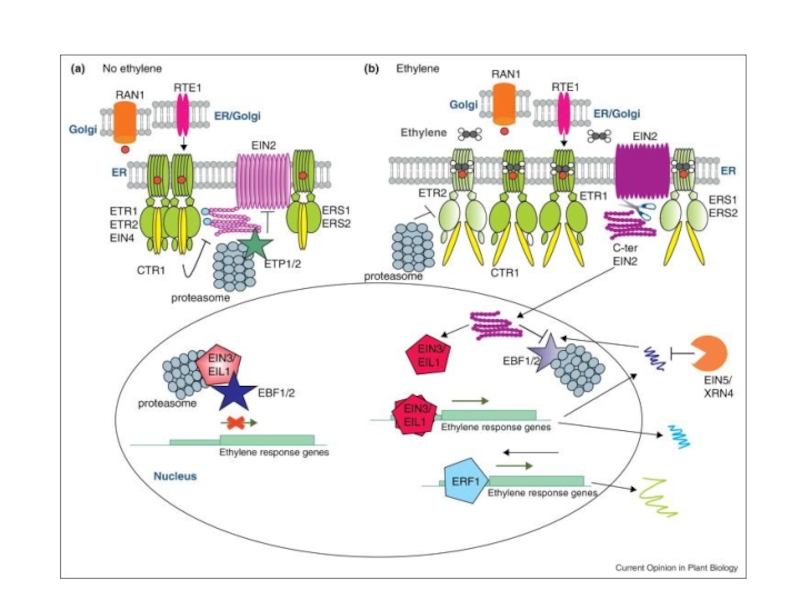
Слайд 23Предполагаемая схема действия МАР-киназного каскада в этиленовом сигналинге
При отсутствии этилена:
Рецепторы активируют
CTR1 инактивирует нижележащие компоненты МАР-киназного каскада: МАРКК (МКК9 Arabidopsis) и МАРК (MPK3/6 Arabidopsis)
В присутствии этилена:
Инактивация CTR1
Активация МКК9 и МРК3/6
МКК9 и МРК3/6 стабилизируют EIN3 ?
Запуск ответа на этилен
PA
Stress response

Слайд 24Белок EIN2 - центральный регулятор этиленового сигналинга
Nramp-like
(не важен для функции)
(важен для функции)
ЭР

Слайд 25Взаимодействие EIN2 с CTR1 и рецепторами этилена
PNAS: November 2012
ETR
CTR1
EIN2
EIN3,
EIL1
Ответ на этилен
ETP1/2

Слайд 26Этилен-зависимые транскрипционные факторы EIN3 и EIL1
Позитивные регуляторы этиленового сигналлинга
Регуляторы транскрипции, локализуются
Сверхэкспрессия генов семейства EIN3 приводит к конститутивному этиленовому ответу
Короткоживущие белки (время полужизни – 30 мин)
Интенсивность ответа на этилен регулируется на уровне стабильности EIN3 белков (но не на уровне экспрессии EIN3 генов)
Индуцируют экспрессию гена ERF1 (Ethylene Response Factor)
EIN3-OE


Слайд 28Регуляция стабильности белков EIN3
Стабилизаторы EIN3: MAPK (фосфорилирование Т174), EIN2 (белок-белковые взаимодействия),
Дестабилизаторы EIN3: CTR1 (фосфорилирование Т592), EBF (убиквитинирование)
, EIN2 (белок-белковые взаимодействия), EIN5 (деградация транскриптов EBF)")
Слайд 29Регуляция стабильности белков EIN3
Белки EIN3 подвергаются протеасомо-зависимой деградации
Связывание белков EIN3
Связывание EIN3 с EBF происходит при фосфорилировании EIN3 белков по консервативному остатку Thr-592, которое осуществляет киназа CTR1
Экспрессия гена EBF2 позитивно регулируется этиленом
Противоположный процесс – стабилизация EIN3 белков
Этилен-зависимая стабилизация белков EIN3 - при фосфорилировании по остатку Thr-174 через МАР-киназный каскад, включающий в себя киназы МКК9 и МРК3/6 (инактивируется CTR1)
Этилен стимулирует накопление EIN3 белков даже у нулевых мутантов ctr1 ? возможен CTR1-независимый путь регуляции (например, через белок EIN2)

Слайд 30
EIN3
EBF genes, etc.
EBF mRNA
EBF proteins
Взаимодействие EIN3-EBF - негативная обратная связь в
MAPK

Слайд 31Рибонуклеаза EIN5/XRN4 регулирует уровень транскриптов EBF1/2
Продукт гена EIN5 был идентифицирован как
Участвует в деградации транскриптов EBF1/2 ? позитивная регуляция этиленового сигналинга
У мутанта ein5 накапливаются белки EBF1/2 ? снижение уровня белка EIN3 ? репрессия ответа на этилен
upstream
signaling

Слайд 32Факты: Сверхэкспрессия 3’UTR генов EBF вызывает нечувствительность к этилену (эффект сильнее,
Механизм: EIN2 взаимодействует с polyU мотивом в 3’UTR транскрипта EBF1, 2, а также с EIN5 и белками PAB. В результате EBF транскрипты не транслируются, образуя РНК-белковые комплексы P-bodies (processing-bodies)
EIN2 репрессирует трансляцию EBF транскриптов

Слайд 33EIN2 репрессирует трансляцию EBF транскриптов
полиU-шпильки в 3’UTR транскриптов EBF
С-концевой домен
Участники:
Экзорибонуклеаза EIN5
Белки РАВ, участвующие в формировании P-bodies всех эукариот



Слайд 36Транскрипционные факторы ERF
Транскрипционные факторы EIN3 и EIL связываются с последовательностью EBS
ERF – семейство АР2-подобных транскрипционных факторов
ERF белки связываются со специфической последовательностью (ERE -Ethylene Response Element; the GCC box)в промоторах этилен-регулируемых генов
EIN5/XRN4
MAPK
Контроль развития
ETP1/2
EIN2
 в промоторе")

Слайд 38ERF-активаторы и ERF-репрессоры
Содержат мотивы EDLL (Глн (E), Асп (D), Лей (L)
Рекрутируют в промотор гена мишени РНК-полимеразу II и гистон-ацетилтрансферазы (НАТ)
Содержат мотивы EAR (ERF-associated Amphiphilic Repression)
Рекрутируют в промотор гена мишени белки-корепрессоры и гистон-деацетилазы (НDА)
AtERF3, 4
AtERF1, 2, 5, 6
, Асп (D), Лей (L) (L))Рекрутируют в промотор гена")
Слайд 39ERFs
Абиотический стресс
(поранение, высокая температура, высыхание)
LEA ? дегидрины
GST ? глутатион-S-трансферазы
Биотический стресс
(поражение
Гены PR белков (pathogene-related): хитиназы, бета-1,3-глюканазы ит.д.
Созревание плодов
Ripening genes:
Е3 (томат) ? пектиназа;
Целлюлазы, экспансины
Старение
Senescence-associated genes (SAG):
Гены протеаз, РНКаз;
Гены ТФ WRKY
LEA ? дегидриныGST ? глутатион-S-трансферазыБиотический стресс (поражение патогенами)Гены PR белков (pathogene-related):")
Слайд 40Некоторые ERF - центральные регуляторы защиты от некротрофных патогенов
Индуцируются в ответ
Основная мишень – гены PR-белков

")
Слайд 42Фитогормоны и системная устойчивость растений
1. ISR (Induced Systemic Resistance)
При
Основные медиаторы – этилен и жасмонаты
2. SAR (Systemic Aquired Resistance)
При взаимодействии с патогенными микроорганизмами
Основной медиатор – салициловая кислота
ethylene response
etr1, ein2
JA response
jar1, coi1
При взаимодействии с непатогенными микроорганизмамиОсновные медиаторы")
Слайд 43Газообразные гормоны – этилен и метил-жасмонат могут выступать в роли вторичных

Слайд 44
Местный (HR) и системный (SAR) иммунный ответ у растений
1
2
3
4
5
PAMPs (pathogen-associated
рецепторы
иммунитет
местный системный
Avr
R
1 – первичная инфекция и синтез сигнальных молекул иммунитета
2 – резкое повышение концентрации салициловой кислоты в месте инфекции – один из механизмов ПКС при НR
3 – развитие системного иммунитета в соседних тканях: более низкая концентрация салициловой кислоты, чем в тканях, подвергающихся НR, ТФ NPR1 активирует транскрицпию генов, кодирующих PR белки
4 – PR белки убивают патогенов, устойчивость к повторному инфицированию
5 – «память иммунитета» за счет модификации гистонов в «генах устойчивости»
 и системный (SAR) иммунный ответ у растений 12345PAMPs (pathogen-associated molecular patterns)рецепторыиммунитетместный системныйAvrR1")
 and Effector-trigegred (ETI) plant immunity: 2 барьера защиты")
Слайд 46PAMP-triggered (PTI) and Effector-trigegred (ETI)
plant immunity: 2 барьера защиты
“Gene
 and Effector-trigegred (ETI) plant immunity: 2 барьера защиты “Gene for gene”")
Слайд 47ТФ NPR1 (nonexpresser of PR genes 1) –
Центральный регулятор иммунитета
Может существовать в виде олигомеров (неактивен) или мономеров (активен), взаимодействовать с репрессорами (NIMIN, WRKY-репрессоры) и активаторами (WRKY-активаторы) транскрипции
 – Центральный регулятор иммунитета растений (SAR)Может существовать в")

Слайд 49Климактерические и неклимактерические плоды
НЕклимактерические – пик интенсивности дыхания при старении
Виноград
Климактерические –
этилен
дыхание плодов

Слайд 50Роль этилена и АБК в созревании плодов
Регулируют одни и те же
Созревание климактерических плодов (томат) – центральная роль этилена
Созревание НЕклимактерических плодов (клубника) – центральная роль АБК

Слайд 51В процессе созревания климактерических плодов повышается биосинтез этилена
ТФ RIN (Ripening Inhibitor)
Синтез этилена
«Размягчение» клеточной стенки
Синтез моносахаридов
Синтез каротиноидов
 и CNR (Colorless Non-Ripening)Синтез")
Слайд 52Роль этилена в старении листьев и созревании плодов
Старение: разрушение хлорофилла, прекращение

Слайд 53Внешние факторы не вызывают старения
Внешние факторы могут вызвать старение
Старение независимо от
Контроль старения листьев в зависимости от «внешних» и «внутренних» причин

Слайд 54Взаимодействие гормонов и ТФ в регуляции возрастозависимого старения листьев
Гормоны «за старение»:
Гормоны «против старения»: CK, BR, GA
Этилен ORE1
АБК NAP
miR164
SAGs
Senescence-Associated Genes
ТФ семейства NAC

Слайд 55Взаимодействие гормонов и ТФ в регуляции старения листьев под воздействием внешних
Этилен
старение в темноте
старение при механических повреждениях
АБК
старение под влиянием дегидратации и засоления

Слайд 56SAG (senescence-associated genes): ТФ семейств NAC, WRKY, AP2
Mишени EIN3 – гены
Этилен и старение листьев
: ТФ семейств NAC, WRKY, AP2Mишени EIN3 – гены OREASA1 и NAP1 –")
Слайд 57Гомологи ORE1 есть у всех наземных растений, гомологи miR164 – у
Синтезировать этилен впервые начали сосудистые растения