- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
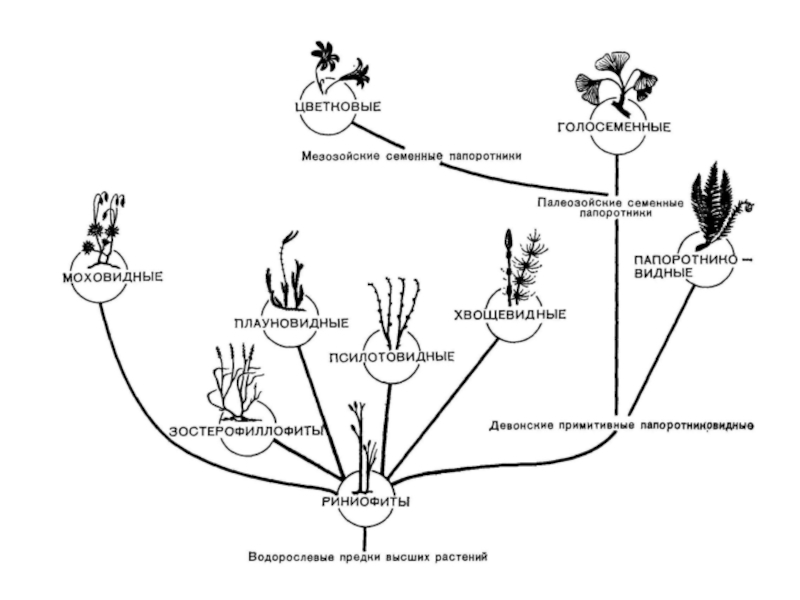
Диплоидная линия эволюции презентация
Содержание
- 1. Диплоидная линия эволюции
- 2. Все высшие растения кроме мхов
- 4. Эволюция жизненного цикла У всех высших растений
- 5. Споровые в наземных условия Приспособления наземных растений
- 6. Массовое производство спор в наземных условиях В
- 7. Расчленение спорофита Увеличение размеров спорофита неизбежно сопровождается
- 8. Появление проводящей системы Увеличение размеров и возрастающее расчленение спорофита сопровождается появлением сложной проводящей системы
- 9. Водное питание в наземных условиях Решение проблем
- 10. Стелярное строение Флоэма и ксилема в центральном
- 11. Стелы разных групп споровых Наиболее примитивным типом
- 12. Эволюция стелы
- 13. Возникновение листьев В результате дальнейшего морфологического расчленения
- 14. Теломные листья Листья большинства споровых и
- 15. Эволюция теломных листьев Теломные листья с самого
- 16. Происхождение листьев
- 18. Теломная теория
- 19. Эволюция ветвления Исходной формой ветвления спорофитов было
- 20. Эволюция ветвления
- 21. Функции листьев у споровых В отличие от
- 22. Разделение функций листьев В процессе эволюции происходило
- 23. Два типа листьев страусопера Трофофиллы Спорофиллы
- 24. побеги со спорофиллами называются стробилами Стробилы споровых
- 25. Среди представителей диплоидной линии эволюции имеются равноспоровые
- 26. Разноспоровые растения Споры разноспоровых растений прорастают в
- 27. Генеральная линия эволюции Таким образом, разноспоровость всегда
- 28. Отделы высших споровых диплоидной линии эволюции
- 30. Риниофиты (RHYNIOPHYTA) или псилофитовидные Впервые они
- 31. Псилофит – голорос первичный (Psilophyton princeps)
- 32. Находка риниофитов Растение было названо Psilophyton princeps
- 33. Представители риниофитов Новые находки получили названия риния (Rhynia) и хорнеофит (Horneophyton).
- 34. Представители риниофитов Позже были найдены другие представители риниофитов: Cooksonia, Yarravia, Trimerophyton и др.).
- 35. Появление синангиев Ярравия
- 36. Строение риниофитов Спорофит риниофитов имел протостелическое строение,

Слайд 2
Все высшие растения кроме мхов
относят к диплоидной эволюционной

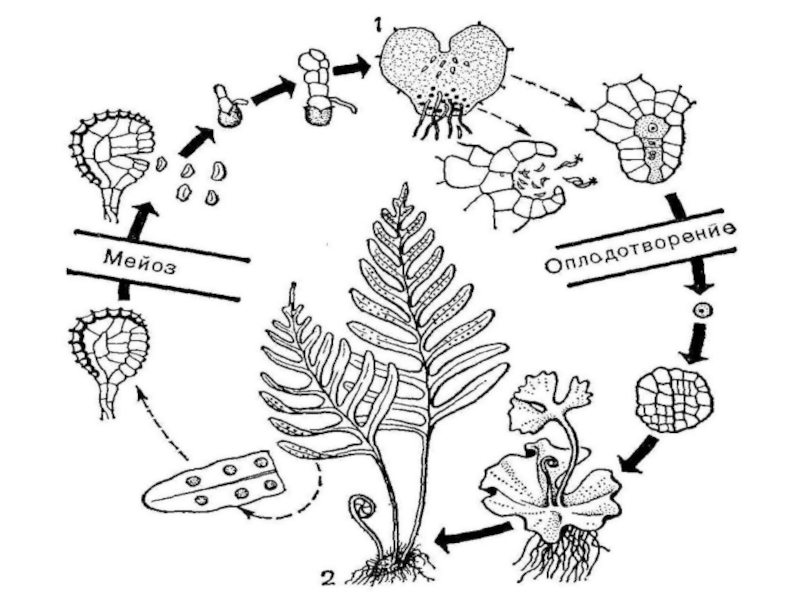
Слайд 4Эволюция жизненного цикла
У всех высших растений спорофит − зеленое автотрофное растение,

Слайд 5Споровые в наземных условия
Приспособления наземных растений выразились в возрастающем расхождении спорофита
появление защищенных оболочкой многоклеточных половых органов, глубокая морфологическая дифференциация спорофита и возникновение многоклеточных спорангиев, развитие проводящей и механической систем тканей, появление эпидермы, кутикулы и устьиц.

Слайд 6Массовое производство спор в наземных условиях
В наземных условиях большая часть спор
Производство большого количества спор требует накопления достаточного количества органического вещества, что неизменно приводит к увеличению размеров самого спорофита и расширению его фотосинтезирующей поверхности.

Слайд 7Расчленение спорофита
Увеличение размеров спорофита неизбежно сопровождается его внешним и внутренним расчленением,

Слайд 8Появление проводящей системы
Увеличение размеров и возрастающее расчленение спорофита сопровождается появлением сложной

Слайд 9Водное питание в наземных условиях
Решение проблем водного режима привело к образованию
У семенных растений в процессе эволюции она пополнилась более крупными сосудами − трахеями разного типа. Органические вещества передвигались по особым ситовидным трубкам − элементам флоэмы.

Слайд 10Стелярное строение
Флоэма и ксилема в центральном цилиндре различных групп высших растений

Слайд 11Стелы разных групп споровых
Наиболее примитивным типом стелы является протостела риниофитов и


Слайд 13Возникновение листьев
В результате дальнейшего морфологического расчленения спорофита возникли специальные органы фотосинтеза
У всех плауновидных они возникли в качестве выростов (энациев) на осевых органах.
Это так называемые энационные листья

Слайд 14Теломные листья
Листья большинства споровых и семенных растений возникли в результате уплощения

Слайд 15Эволюция теломных листьев
Теломные листья с самого начала были спороносными, выполняя как
В процессе дальнейшей эволюции происходило постепенное пространственное разделение этих функций.




Слайд 19Эволюция ветвления
Исходной формой ветвления спорофитов было равнодихотомичес-кое ( риниофиты, плауны, псилот),
Дальнейшим шагом в эволюции ветвления стало дихоподиальное ветвление, которое постепенно перешло в моноподиальное (хвойные) и, наконец, в процессе эволюции возникло симподиальное ветвление, широко распространенное у цветковых растений.
, после чего растения выработали")

Слайд 21Функции листьев у споровых
В отличие от безлистных риниофитов на листьях высших
Теломные листья в отличие от энационных с самого начала были спороносными (фертильными). Такие спороносные листья, или спорофиллы, исполняли сначала функцию фотосинтеза и бесполого размножения

Слайд 22Разделение функций листьев
В процессе эволюции происходило разделение этих функций. Так у



Слайд 25Среди представителей диплоидной линии эволюции имеются равноспоровые и разноспоровые растения
У
У разноспоровых на спорофите образуются спорангии двух типов: мегаспорангии с мегаспороами и микроспорангии с микроспорами.

Слайд 26Разноспоровые растения
Споры разноспоровых растений прорастают в однополые, сильно редуцированные микроскопические гаметофиты,
Особенно сильно редуцирован мужской гаметофит, нередко представляющий всего одну − две ризоидальные клетки

Слайд 27Генеральная линия эволюции
Таким образом, разноспоровость всегда сопровождается сильнейшей редукцией гаметофита.
Редукция гаметофитов,


Слайд 30Риниофиты (RHYNIOPHYTA) или псилофитовидные
Впервые они были найдены в 1859 г. канадским
 или псилофитовидныеВпервые они были найдены в 1859 г. канадским геологом Дж. Досоном в")
")
Слайд 32Находка риниофитов
Растение было названо Psilophyton princeps − голорос первичный. Необычное растение
Повторно риниофиты были обнаружены в 1912 г. в местечке Райни в Шотландии в пластах раннедевонского (415 млн. лет назад) кремнистого черта.

Слайд 33Представители риниофитов
Новые находки получили названия риния (Rhynia) и хорнеофит (Horneophyton).
 и хорнеофит (Horneophyton).")
Слайд 34Представители риниофитов
Позже были найдены другие представители риниофитов: Cooksonia, Yarravia, Trimerophyton и
.")

Слайд 36Строение риниофитов
Спорофит риниофитов имел протостелическое строение, а механические ткани и камбий






