- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Детермінація пола презентация
Содержание
- 2. Гаплоидный и диплоидный жизненный цикл.
- 4. Бделлоидные коловратки представляют собой удивительное исключение. Это
- 5. Англійські та італійськи вчені повідомили про дивне
- 6. ПАРТЕНОГЕНЕЗ 1.ЭНДОМІТОЗ: zw4zw zw+3 пт(zw) все
- 12. Вильям Хамильтон: компьютерная модель полового/бесполого размножения и
- 13. Теория красной королевы («черного ферзя») Ли Ван
- 14. Теория красной королевы Только рекомбинация генов при
- 15. Упрощенная схема пространственного распределения фрактофузусов.Nature2015 Fractofusus —
- 16. The 385 million-year-old placoderm fish Microbrachius dicki
- 18. Типы гамет у хламидомонад: изогамия, анизогамия, оогамия
- 19. СПОСОБИ ВИЗНАЧЕННЯ СТАТІ 1.СІНГАМНИЙ (у
- 20. Кореляція між співвідношенням статей F / (F
- 21. Залежне від температури (інкубації яєць) визначення статі
- 22. Температурно-залежна детермінація статі у ящірок, черепах і крокодилів
- 23. У эхиуриды Bonellia (организма, обитающего на морском
- 24. У улитки- блюдечка Crepidula fornicata, новая особь,
- 25. У риб поряд з раздільностатевими видами є
- 26. Зміни структури гонад гермафродитної риби Sparus aurata
- 27. Хромосомне визначення статі. У тварин існують
- 28. Походження статевих хромосом Порівняльний аналіз статевих хромосом
- 29. У різних груп хребетних тварин стать визначається
- 30. Існують два основних правила визначення статі у
- 31. Появление и происхождение гинандроморфных птиц
- 32. Гинандроморфізм поєднання у особини роздільностатевого виду чоловічих
- 33. Гинандроморфы дрозофилы и мотылька, образовавшиеся в результате
- 34. Тільце Барра в клітині. У самиць
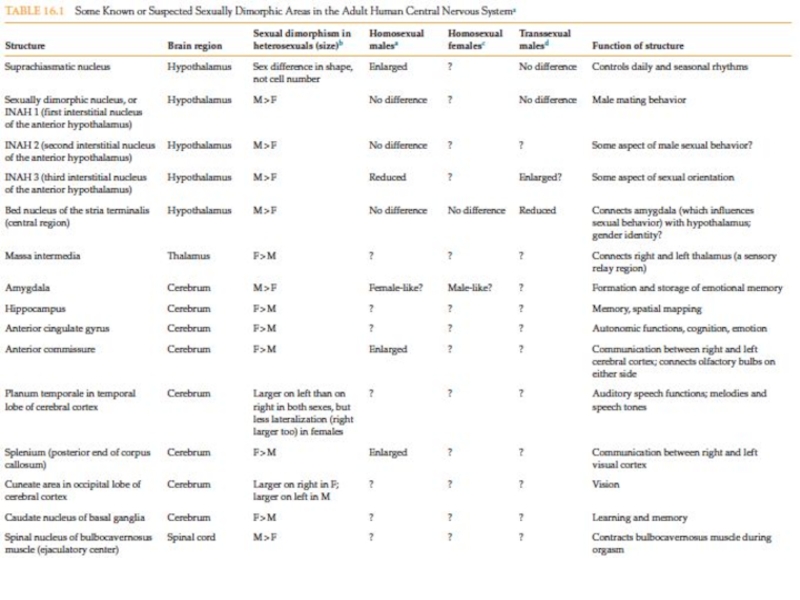
- 36. Первичная и вторичная детерминация пола у млекопитающих
- 37. Бессмертная Turritopsis nutricula. Вверху : жизненый цикл обычной сцифоидной медузы. Слева: трансдифференциация бессмертной нутрикулы
- 39. Первичные половые клетки (ППК) у млекопитающих, производные
- 40. Маркёрными белками ППК млекопитающих являются фактор транскрипции Oct4 и щелочная фосфатаза
- 41. Після міграції з ектодерми первинні зародкові клітини
- 42. Mouse E9.0 Primordial Germ Cell Migration
- 43. Mouse E9.5 Primordial Germ Cell Migration
- 44. MMMouse E10.5 Primordial Germ Cell Migration
- 45. Зачаток гонадного валику на 5-му тижні розвитку.
- 46. Розвиток сім’яних канальців в зачатку сім’яника. 1.
- 47. Дифференцировка гонад человека (начало)
- 49. Дифференцировка бипотенциальных гонад и образование половых структур из Вольфова и Мюллерова протоков
- 50. Дифференцировка гонад человека (продолжение)
- 51. Дифференцировка гонад человека (окончание)
- 52. При анализе Y-хромосомы выяснилось, что гипотетический фактор,
- 54. Мышь с половым генотипом XX, трансгенная по
- 55. Проникновение (миграция) мезонефротических клеток (любого генотипа: XY
- 56. Экспрессия гена Sry приводит к активации генов
- 58. Роль антимюллерова гормона в дегенерации мюллерова протока
- 59. Выявление гена DAX1 на X-хромосоме, ответственного за
- 62. Тестостерон и 5 α-дигидротестостерон - андрогены. Тестостерон
- 63. Районы мужской половой системы, формирующиеся под контролем тестостерона и дидротестостерона.
- 64. Мужские и женские стероидные гормоны. Эстрогены (включая
- 65. Виды женских половых гормонов и их функции
- 66. Эволюционные связи между генами, ответственными за детерминацию
- 67. Сколько раз в эволюции возникали механизмы детерминации
- 68. Пути биосинтеза мужских и женских стероидных гормонов у позвоночных (на примере млекопитающих)
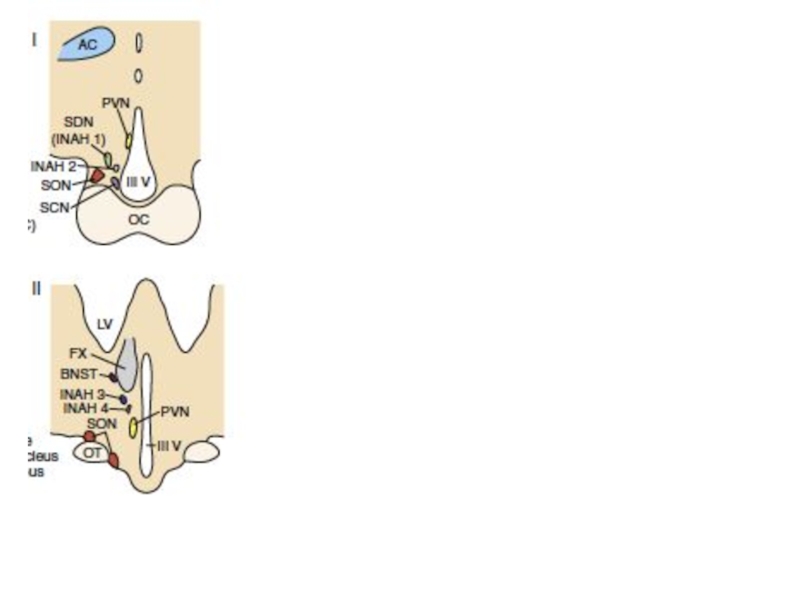
- 69. A, arcuate nucleus of the hypothalamus (GnRH


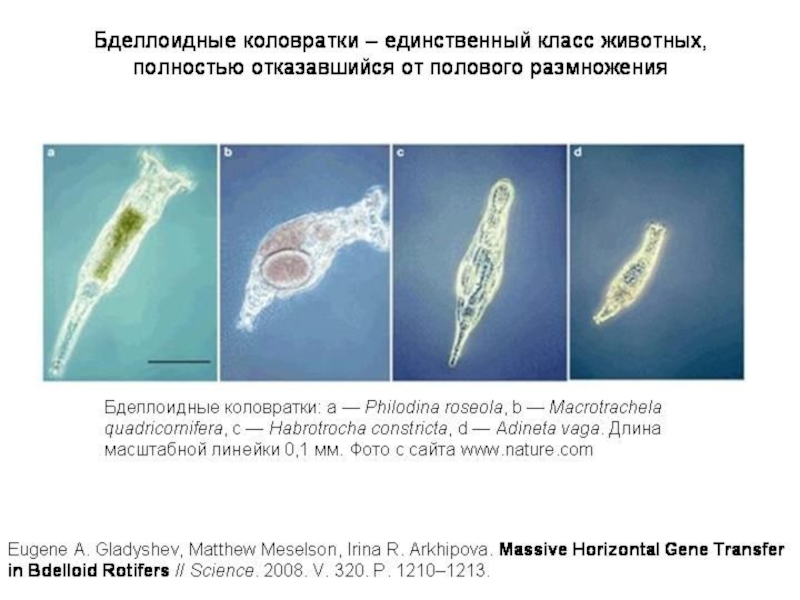
Слайд 4Бделлоидные коловратки представляют собой удивительное исключение. Это целый класс животных (включающий
Бделлоидные коловратки — главный камень преткновения для всех теоретиков, пытающихся объяснить биологический смысл полового размножения. Какое ни придумай объяснение, сразу же возникает «проклятый» вопрос: если половое размножение такое полезное, как же бделлоидные коловратки без него обходятся? И если бделлоидные коловратки научились без него обходиться, почему другие животные не пошли по тому же пути?
, которые")
Слайд 5Англійські та італійськи вчені повідомили про дивне відкриття – на відміну
Скельні ящерки
Комодський варан

Слайд 6ПАРТЕНОГЕНЕЗ
1.ЭНДОМІТОЗ: zw4zw zw+3 пт(zw) все самки
2.ВИПАДІННЯ МЕЙОЗУ (обидва ділення
3.ПРИДУШЕННЯ 1 ПОДІЛУ ДОЗРІВАННЯ (прередукція): все zw (самиці)
4. ПРИДУШЕННЯ 2 РОЗПОДІЛУ ДОЗРІВАННЯ (постредукція): все zz (самці), ww – let
5. НОРМАЛЬНИЙ МЕЙОЗ, ЗЛИТТЯ БЛАСТОМЕРІВ ПІСЛЯ 1 РОЗПОДІЛУ ДРОБЛЕННЯ: все zz (самці), ww – let
6.НОРМАЛЬНИЙ МЕЙОЗ, ЗЛИТТЯ ЯЙЦЕКЛІТИНИ З ПОЛЯРНИМ ТІЛЬЦЕМ: 4 zw(): 1 zz(): 1 ww(let)
 все самки 2.ВИПАДІННЯ МЕЙОЗУ (обидва ділення экваційні): все zw (самиці)")
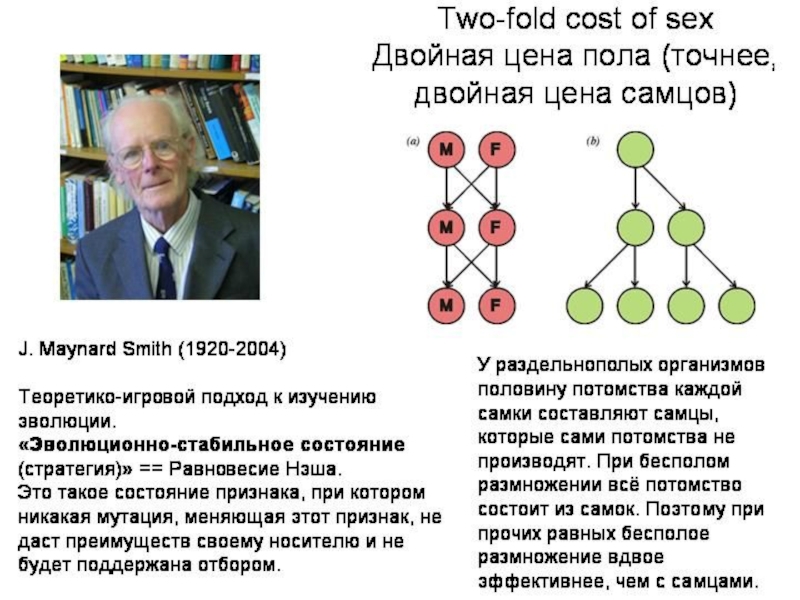
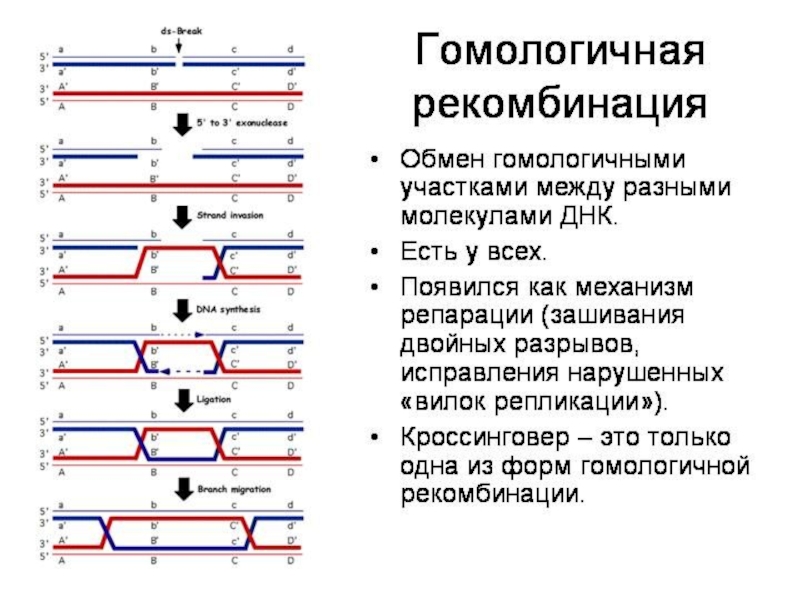

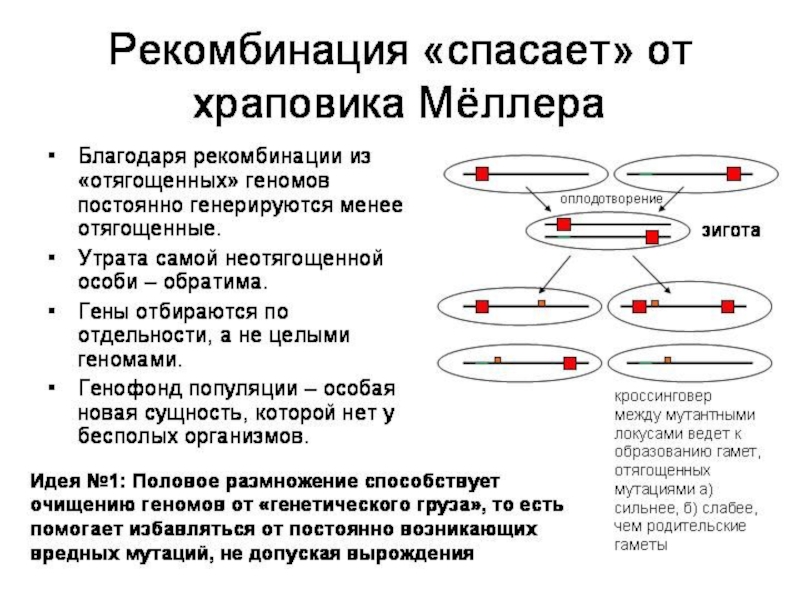
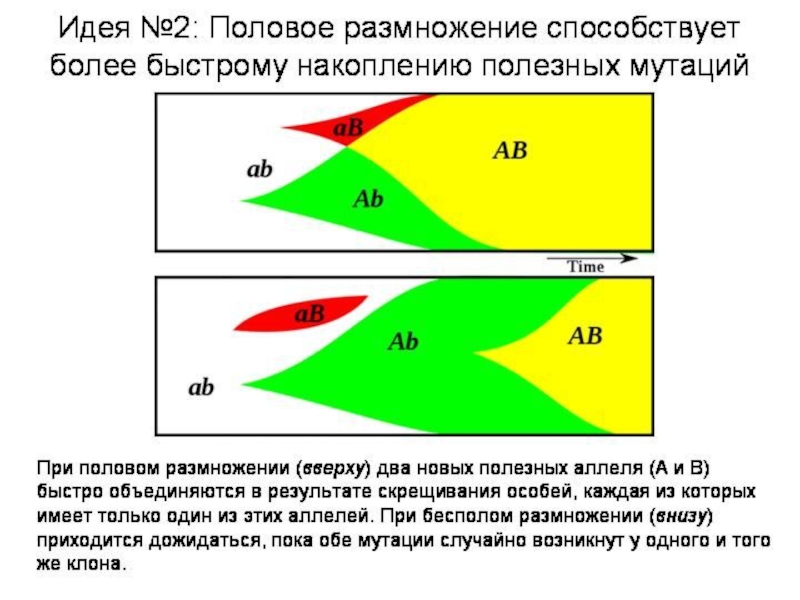
Слайд 12Вильям Хамильтон: компьютерная модель полового/бесполого размножения и болезни
Двуполые организмы выигрывают только

Слайд 13Теория красной королевы («черного ферзя») Ли Ван Вален
- У нас, -
- Какая медлительная страна! - сказала Королева. - Ну, а здесь, знаешь ли, приходится бежать со всех ног, чтобы только остаться на том же месте! Если же хочешь попасть в другое место, тогда нужно бежать по меньшей мере вдвое быстрее!
 Ли Ван Вален- У нас, - сказала Алиса, с трудом")
Слайд 14Теория красной королевы
Только рекомбинация генов при мейозе и ауткроссинг генов при
Половое размножение – это единственный шанс выжить для популяции медленно размножающихся макроорганизмов в условиях постоянного давления со стороны быстро размножающихся и мутирующих микроорганизмов

Слайд 15Упрощенная схема пространственного распределения фрактофузусов.Nature2015
Fractofusus — организмы эдиакарского периода из группы

Слайд 16The 385 million-year-old placoderm fish Microbrachius dicki from Scotland shows the



Слайд 19СПОСОБИ ВИЗНАЧЕННЯ СТАТІ
1.СІНГАМНИЙ (у ссавців) - за допомогою статевих хромосом,
2.ПРОГАМНИЙ (у археаннелід, попелиць, дафній) - ще до запліднення, стать визначається типом відкладених яєць (розмір, тестостерон та інш)
3.ЕПІГАМНИЙ (у деяких молюсків, риб та рептилій) - стать визначається умовами середовища
 - за допомогою статевих хромосом, стать визначається в момент")
Слайд 20Кореляція між співвідношенням статей F / (F + M) і температурою
 і температурою у риби Menidia menidia")
Слайд 21Залежне від температури (інкубації яєць) визначення статі у трьох представників класу
 визначення статі у трьох представників класу плазунів: грифова (Macroclemis) і")

Слайд 23У эхиуриды Bonellia (организма, обитающего на морском каменистом грунте) формирование пола
 формирование пола происходит у личинки в")
Слайд 24У улитки- блюдечка Crepidula fornicata, новая особь, располагающаяся в колонии сверху

Слайд 25У риб поряд з раздільностатевими видами є і гермафродити Гермафродити можуть


Слайд 27Хромосомне визначення статі. У тварин існують статеві хромосоми, тому стать визначається в
zz top
самцы

Слайд 28Походження статевих хромосом
Порівняльний аналіз статевих хромосом в різних таксонах дозволяє виділити

Слайд 29У різних груп хребетних тварин стать визначається по-різному : червоними кругами

Слайд 30Існують два основних правила визначення статі у ссавців:
перше з них було
перше правило: видалення гонадних валиків до формування гонад приводить до розвитку всіх ембріонів як самок;
друге правило : Y-хромосома несе генетичну інформацію, що потрібна для визначення статі у самців.
Комбінацію двох правил називають принципом росту


Слайд 32Гинандроморфізм поєднання у особини роздільностатевого виду чоловічих і жіночих ознак.
Гінандроморфи діляться
Білатеральні: одна поздовжня половина тіла чоловічої статі, інша - жіночої;
Передньо-задні: передня частина однієї статі, задня - іншої;
Мозаїчні: у яких перемежовуються ділянки тіла, що несуть ознаки різних статей.

Слайд 33Гинандроморфы дрозофилы и мотылька, образовавшиеся в результате утраты в раннем развитии

Слайд 34Тільце Барра в клітині.
У самиць ссавців, гетерозиготніих за будь-якою ознакою,
Лайонізація (Інактивация Х-хромосоми).

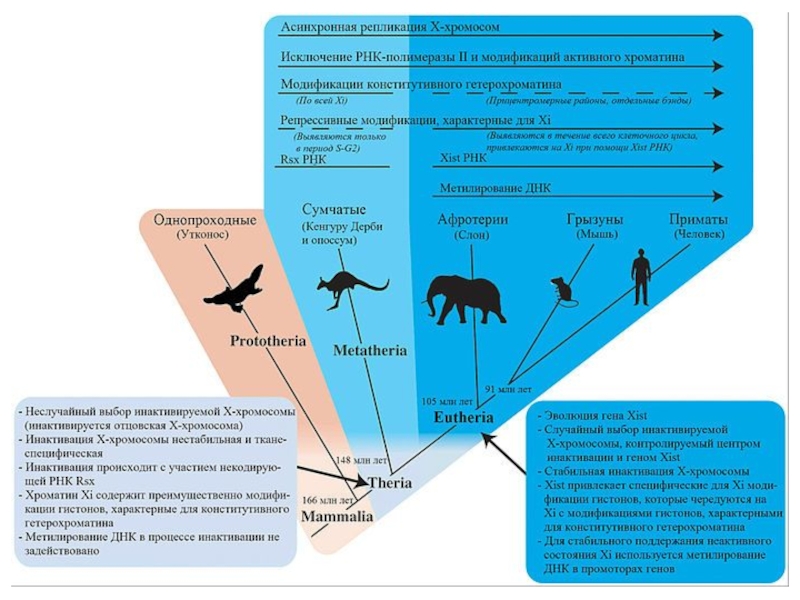
Слайд 36Первичная и вторичная детерминация пола у млекопитающих Первичная детерминация пола – это

Слайд 37Бессмертная Turritopsis nutricula.
Вверху : жизненый цикл обычной сцифоидной медузы.
Слева: трансдифференциация

Слайд 39Первичные половые клетки (ППК) у млекопитающих, производные клеток внутренней массы, выявляются
 у млекопитающих, производные клеток внутренней массы, выявляются в пограничном районе желточного")

Слайд 41Після міграції з ектодерми первинні зародкові клітини тепер розташовані серед клітин
Первинні статеві клітини переміщуються вздовж стінки жовткового мішку, жовткової і спинної брижі в гонадний валик. Досягають цю структуру на 6-му тижні. 1.Пряма кишка;2. Жовтковий мішок; 3. Алонтоіс; 4. Нефрогенний тяж; 5. Гонадний валик (зелений); 6. Первинні статеві клітини (червоні точки) ; 7. Серце.




Слайд 45Зачаток гонадного валику на 5-му тижні розвитку. Великий малюнок : формування

Слайд 46Розвиток сім’яних канальців в зачатку сім’яника. 1. Вольфів канал; 2-первинні статеві
Розвиток яєчнику. Зародкові тяжі дегенерують у віддаленому від стінки регіоні зачатку гонади регіоні. Внаслідок проліферації епітелію порожнини тіла зародкові тяжі, які не втратили з нею контакт залишаються. 1.-Вольфів канал; 2 - первинні статеві клітини; 3 – черевна порожнина; 4 – аорта; 5 – канальці мезонефросу; 6 – дегенеруючи гонадні тяжі;7- епітелій порожнини тіла; 8 – епітелій кишечнику;9 – брижейка; 10- зачаток каналу Мюллера; 11 – Нефрон мезонефросу.

")
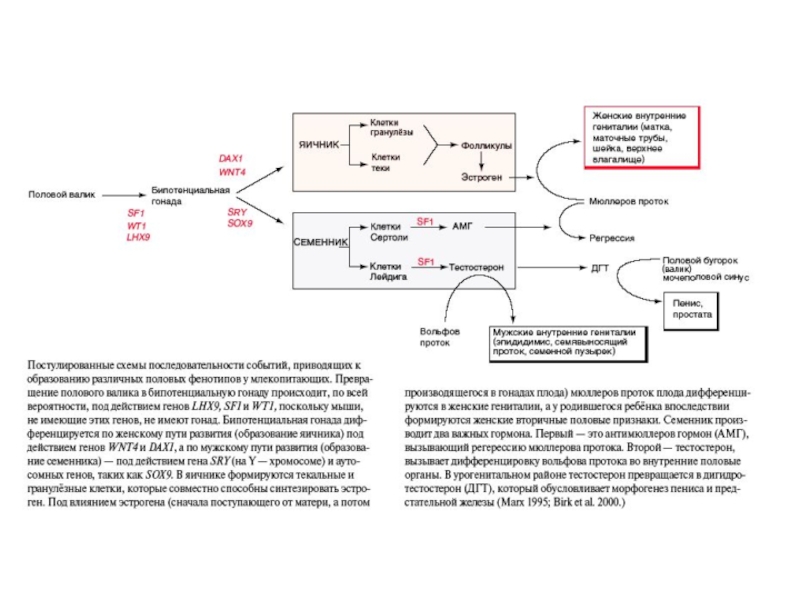
Слайд 49Дифференцировка бипотенциальных гонад и образование половых структур из Вольфова и Мюллерова

")
")
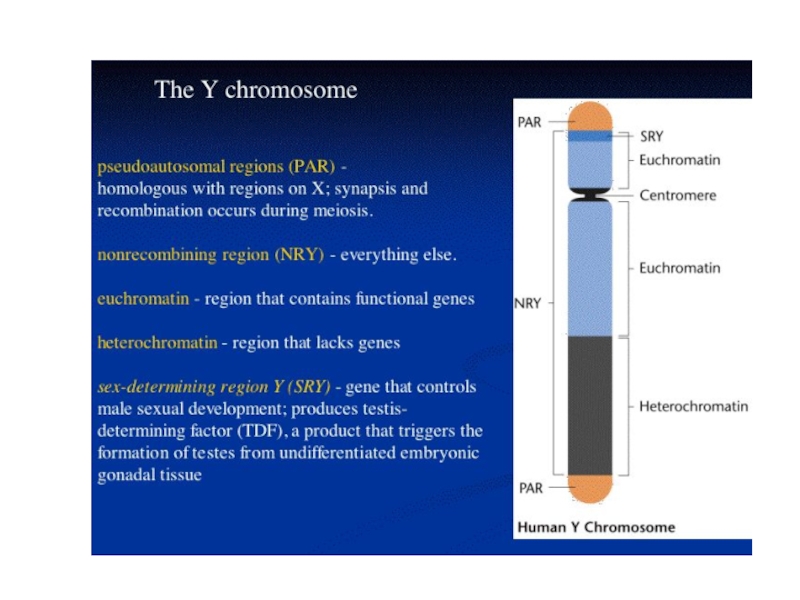
Слайд 52При анализе Y-хромосомы выяснилось, что гипотетический фактор, отвечающий за формирование пола,

Слайд 54Мышь с половым генотипом XX, трансгенная по гену SRY(справа), в норме
, в норме локализованным на Y-хромосоме обладает")
Слайд 55Проникновение (миграция) мезонефротических клеток (любого генотипа: XY или XX) в Sry+
 мезонефротических клеток (любого генотипа: XY или XX) в Sry+ гонадные валики (генотипа XY)")
Слайд 56Экспрессия гена Sry приводит к активации генов SF1 и Sox9 в

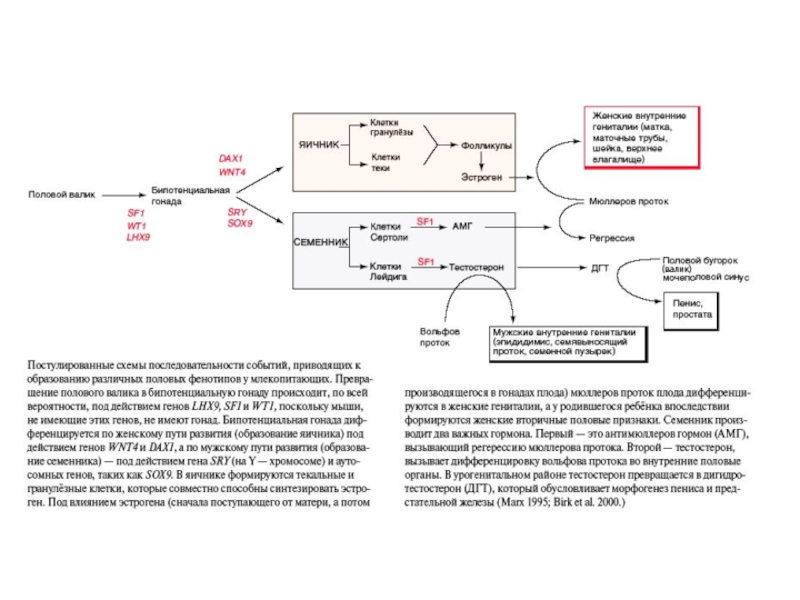
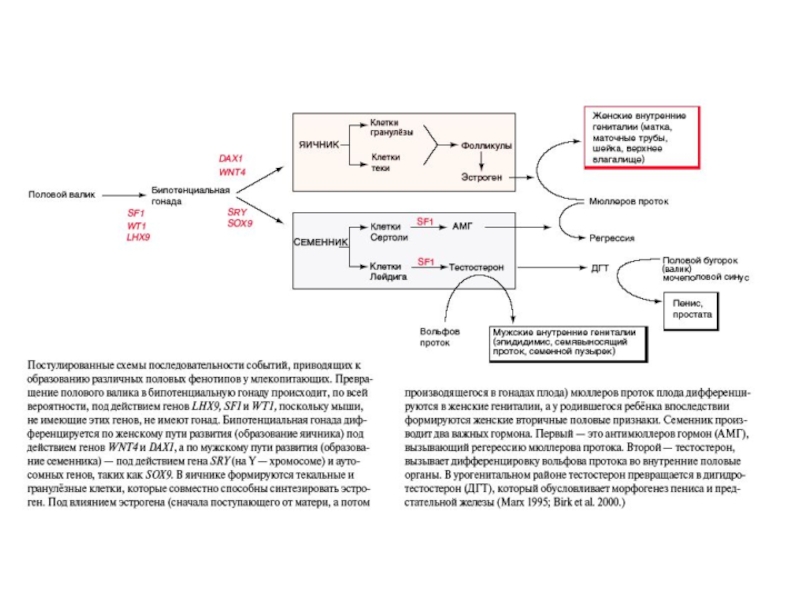
Слайд 58Роль антимюллерова гормона в дегенерации мюллерова протока (плод крысы). Слева -
. Слева - открыты оба протока, справа")
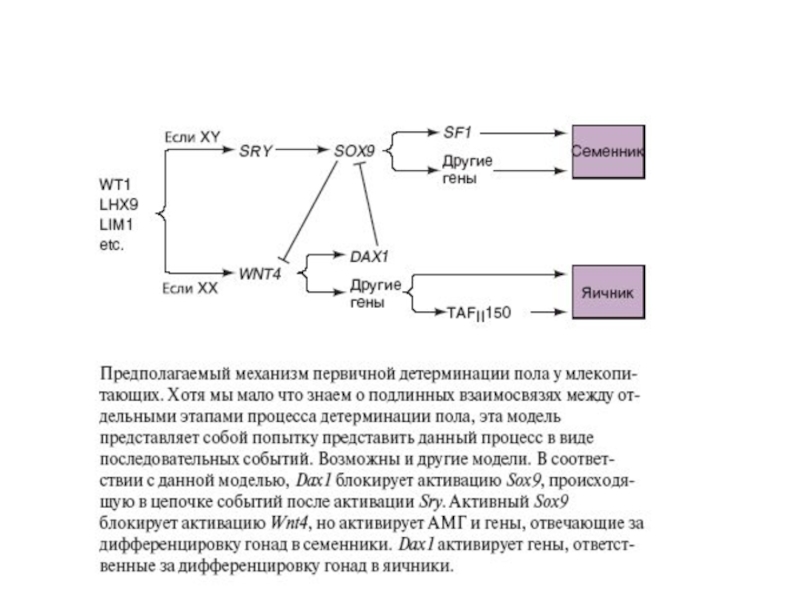
Слайд 59Выявление гена DAX1 на X-хромосоме, ответственного за формирование женского пола. Ген

Слайд 62Тестостерон и 5 α-дигидротестостерон - андрогены. Тестостерон синтезируется в клетках Лейдига

Слайд 63Районы мужской половой системы, формирующиеся под контролем тестостерона и дидротестостерона.

Слайд 64Мужские и женские стероидные гормоны. Эстрогены (включая их аналоги- диэтилстилбестрол и
 важны не")

Слайд 66Эволюционные связи между генами, ответственными за детерминацию пола у разных животных
В

Слайд 67Сколько раз в эволюции возникали механизмы детерминации пола? Гипотеза: Cуществуют две группы позвоночных

Слайд 68Пути биосинтеза мужских и женских стероидных гормонов у позвоночных (на примере
")
Слайд 69A, arcuate nucleus of the hypothalamus (GnRH cells
migrating to here control
pathways of GnRH cells.
Diagram of a section through the head of a human embryo showing
the migration route of GnRH neurons (green dots) from the
nasal cavity to the hypothalamus.