- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Биотехнология растений. Трансгенные растения (часть 1) презентация
Содержание
- 1. Биотехнология растений. Трансгенные растения (часть 1)
- 2. Трансгенные растения-1 Молекулярные механизмы взаимодействия растения
- 3. Нобелевская премия , основоположник 1и2-ой Зеленой революции.
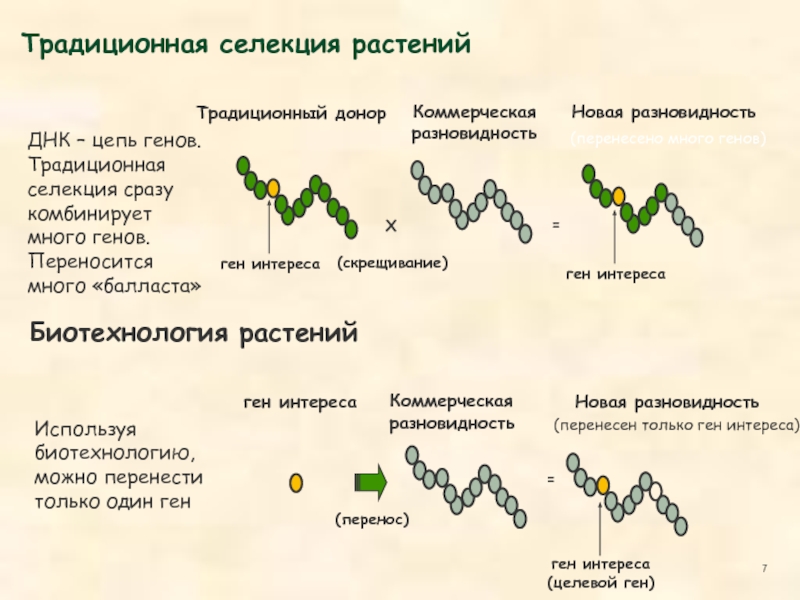
- 4. X Методы традиционной селекции растений 1. Близкородственное
- 5. Slide courtesy of Wayne Parrott, University of Georgia теосинте кукуруза
- 6. Slide courtesy of Wayne Parrott, University of Georgia
- 8. Генетически модифицированный организм (ГМО) - организм, генетический
- 9. 1975г. - группы из компании Монсанто, из
- 10. Значительное ускорение создания сорта (1-3 года против
- 12. Основные культуры трансгенных растений
- 13. Основные направления биотехнологии: производство продуктов питания с
- 14. Векторные системы для переноса генов в растения Вирусы растений Транспозоны Агробактерии
- 15. Схема агробактериальной трансформации
- 16. Геном агробактерий
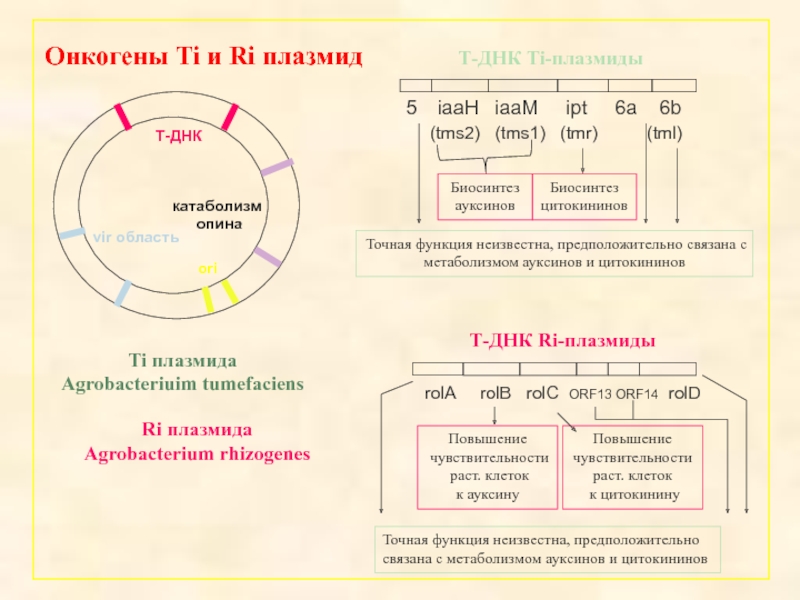
- 17. Октопин Нопалин Агропин Структура Ti-плазмиды и основные опины
- 18. Онкогены A. tumefaciens 5 1. tms-гены
- 19. Онкогены A. tumefaciens 6 2. tmr-ген
- 20. Онкогены A. tumefaciens 7 3. tml-локус(6a,
- 22. Онкогены A. rhizogenes. 8 RolA -
- 23. Онкогены A. rhizogenes. 9 ORF13 –
- 24. Пути биосинтеза основных гормонов в агробактерии
- 25. Сигнальные молекулы
- 26. Опиновая концепция 12
- 27. Интеграция Т-ДНК в геном растения
- 28. Требование к векторным системам Идеальная векторная система
- 29. pBR322 Ti-плазмида + T-ДНК T-ДНК pBR322
- 30. Бинарный вектор Коинтегративный вектор
- 32. Репортерные гены Ген GFP Ген GUS Ген LUX
- 33. Биолюминисценция у Aequoria victoria aequorin Зеленый флюоресцентный белок
- 34. Репортерные гены
- 35. Методы прямого переноса электропорация биобаллистика трансфекция Методы
- 36. Схема баллистической трансформации
- 37. Трансформация картофеля Agrobacterium Образование микроклубней (1
- 38. Трансформация гороха Agrobacterium Спелый горох 96%
- 40. Eugene Nester и Frank White обнаружили в
- 41. N. otophora N.tomentosiformis rolC, ORF13, ORF14 rolC,
- 42. (Suzuki et.al., 2001, 2002) Актов трансформации в
- 43. Expression of the oncogenes (Intrieri M.C., Buiatti M., 2001)
- 44. Скрининг видов на наличие Т-ДНК содержащих последовательностей
- 45. п/кл. Ranunculidae сем. Berberidaceae (1) сем. Ranunculaceae
- 46. В 2006 году сотрудниками лаборатории ГКИР был
- 47. Выявлены последовательности, гомологичные rolB, rolC, ORF13, ORF14
- 48. Анализ района инсерции у разных линарий. У
- 49. Обнаружение явления косупрессии у растений (Alexander

Слайд 2 Трансгенные растения-1
Молекулярные механизмы взаимодействия растения – агробактерии
Векторы на основе агробактерии
Селективные
Методы трансформации
Замолкание (сайленсинг) генов у трансгенных растений

Слайд 3Нобелевская премия , основоположник 1и2-ой Зеленой революции.
Норман Эрнст Борлаг
Норман Борлаг получил
пшеницы с повышенной урожайностью.
Начало «Зеленой Революции» в сельском
хозяйстве.
1963

Слайд 4X
Методы традиционной селекции растений
1. Близкородственное скрещивание (имбридинг)
родительская форма 1
потомство
отбор
2. Отдаленное скрещивание
родительская форма 1
X
родительская форма 1
потомство (гетерозисные гибриды)
отбор
родительская форма 2
3. Мутагенез
родительская форма
потомство (мутанты)
отбор
мутагенез
родительская форма 1потомствоотбор2. Отдаленное скрещивание (аутбридинг)родительская форма 1Xродительская форма")


Слайд 8Генетически модифицированный организм (ГМО) -
организм, генетический материал которого (ДНК) изменен не
 - организм, генетический материал которого (ДНК) изменен не в ходе естественной гибридизации")
Слайд 91975г. - группы из компании Монсанто, из Гентского государственного университета (Бельгия),
1990г. - первое коммерческое применение ГМО в США.
1992г. - в Китае начали промышленно выращивать трансгенный табак, устойчивый к насекомым.
1994г. - в США зарегистрировали первое трансгенное растение, предназначенное для употребления в пищу, - томаты “Флавр-Савр” с замедленным созреванием.
1999г. - получены трансгенные растения более чем 120 видов.
История развития получения трансгенных растений
, из Института растениеводства им.")
Слайд 10Значительное ускорение создания сорта (1-3 года против 10 и более лет).
Избавление
Создание растений с заданными признаками. Традиционная селекция отбирает растения, которые нас устраивают, биотехнология создает растения, которые нам нужны.
Большая возможность контроля целевого гена за счет управления его экспрессией в нужных органах, тканях и в нужное время.
В чём преимущества методов генной инженерии по сравнению с традиционной селекцией?
.Избавление от значительного количества «генетического")
Слайд 11
Уолтер Гилберт
Создание
устойчивостью к гербицидам, вредителям и вирусам.
1980-е годы


Слайд 13Основные направления биотехнологии:
производство продуктов питания с заданными характеристиками;
производство веществ вторичного метаболизма
получение растений с декоративными признаками;
использование трансгенных растений в фундаментальных исследованиях.



Слайд 16
Геном агробактерий
A. rhizogenes
A. tumefaciens
A. vitis
3Мб
3Мб
3Мб
2Мб хромосома II
2Мб хромосома II
2,65Мб
3Мб
450кб
450кб
200кб
200кб
Хромосома I
Хромосома I
Хромосома
pAt
pRi
pTi
pTi
pAt
200кб
2


Слайд 18
Онкогены A. tumefaciens
5
1. tms-гены (iaaM, iaaH)
Триптофан
Индолацетамид
Ауксин
iaaM
iaaH
Гены бактериального происхождения, гомологи
аналогичных генов
фитопатогенов
Pseudomonas savastanoi
iaaH
6b
iaaM
ipt
5
 ТриптофанИндолацетамидАуксинiaaMiaaHГены бактериального происхождения, гомологианалогичных генов у других ауксин-синтезирующих фитопатогенов")
Слайд 19
Онкогены A. tumefaciens
6
2. tmr-ген (ipt)
HMBDP
Зеатинрибозид – 5-МФ
Транс-зеатин
ipt
iaaH
6b
iaaM
ipt
5
 HMBDPЗеатинрибозид – 5-МФТранс-зеатинiptiaaH6biaaMipt5")
Слайд 20
Онкогены A. tumefaciens
7
3. tml-локус(6a, 6b)
6a – опиновая пермеаза
4. ген 5.
Индол-3-лактат
5
6b – онкоген с неизученным механизмом действия, взаимодействует с
некоторыми хозяйскими ТФ.
Триптофан
 6a – опиновая пермеаза4. ген 5. Индол-3-лактат56b – онкоген с")
Слайд 22
Онкогены A. rhizogenes.
8
RolA - есть на всех Ri-плазмидах. Снижает концентрацию ауксина,
RolB – рассматривается как основной онкоген A.
rhizogenes. Повышает чувствительность клеток
к ауксину
RolС –b-гликозидаза. Гидролизует неактивные
цитокинин-гликозиды или влияет на клеточный цикл
через регуляцию метаболизма сахаров.
RolD – орнитин-циклодеаминаза
Набор онкогенов A.rhizogenes не совпадает с набором онкогенов A.tumefaciens и не всегда содержит гены синтеза гормонов.
Пролин
Орнитин
RolD

Слайд 23
Онкогены A. rhizogenes.
9
ORF13 – связывает белок ретинобластомы (Rb). Стимулирует
пролиферацию клеток,
В конечном итоге способствует дедифференцировке трансформированных
клеток и образованию новой меристемы
aux1, aux2 – гены синтеза ауксина, гомологи iaaM и iaaH. Только у агропиновых штаммов.
. Стимулирует пролиферацию клеток, вызывает эктопическую экспрессию KNOX-генов.В")




Слайд 28Требование к векторным системам
Идеальная векторная система на основе Ti-плазмиды должна содержать:
сигналы,
узнаваемый растительными полимеразами промотор;
селективный маркер для селекции трансформированных клеток;
репортерные гены для отбора трансгенных растений;
полилинкер (уникальные сайты рестрикции), в который будет клонирован чужеродный фрагмент ДНК;
не содержать онкогены.

Слайд 29pBR322
Ti-плазмида
+
T-ДНК
T-ДНК
pBR322
Ген растения
Встраивание гена растения в несущественный участок Т-ДНК
pBR322
Agrobacteria
Гомологичная рекомбинация с Ti-плазмидой
Ti-плазмида
ДНК растения
Интеграция Т-ДНК с новым геном в геном растения
Схема конструирования вектора на основе Ti-плазмиды






Слайд 35Методы прямого переноса
электропорация
биобаллистика
трансфекция
Методы непрямого переноса
вирусная трансформация (трансдукция)
агробактериальная трансформация
Методы трансформации
агробактериальная трансформацияМетоды трансформации")

Слайд 37Трансформация картофеля Agrobacterium
Образование микроклубней (1 мес.)
Иннокуляция клубневых дисков A. tumefaciens (15
Регенерация побегов
Укоренение на среде, содержащей Km устойчивых к норфлуразону регенерантов
Селекция на среде, содержащей 100 мг/л Km
Ко-культивация с A. tumefaciens (3 д)
Ночная культура A. tumefaciens
Иннокуляция клубневых дисков A. tumefaciens (15 с)Регенерация побеговУкоренение на среде,")
Слайд 38Трансформация гороха Agrobacterium
Спелый горох
96% этанол, 20 минут
Удаление корней и разделение горошины
Среда B5: + 0,5 мг/л 2-4D 4,5 мг/л BAP
Среда MS: + 5 мг/л BAP 2 мг/л NAA, с Agrobacterium
Среда MS: + 5 мг/л BAP 2 мг/л NAA, 500 мг/л Cf, 5 мг/л Basta
Среда MS: + 5 мг/л BAP 2 мг/л NAA, 500 мг/л Cf, 5 мг/л Азацетидин
Фильтровальная бумага
Вакуум
Агробактериальная суспензия
48 ч
20 д
ПЦР, GUS окрашивание


Слайд 40Eugene Nester и Frank White обнаружили в геноме
нетрансформированного Nicotiana glauca
гомологичные участку плазмиды Agrobacterium rhizogenes.
Nicotiana glauca

Слайд 41N. otophora
N.tomentosiformis
rolC, ORF13, ORF14
rolC, rolB, ORF13, ORF14
N.bigelovii
rolB
rolC
N.acunminata
N.benavidensii
N.gossei
N. suaveolens
N. arentsii
N. tabacum
N. glauca
Примеры

Слайд 42(Suzuki et.al., 2001, 2002)
Актов трансформации в эволюции рода Nicotiana было несколько
Существуют две группы Т-ДНК содержащих видов
Данные на основании сравнения приграничных последовательностей, а также гомологов микимопин синтазы.
Схема дивергенции части рода Nicotiana (черные и белые стрелки показывают предполагаемое инфицирование плазмидой микимопинового типа и неизвестного типа опинов, соответственно. Виды, содержащие Т-ДНК Ri плазмиды микимопинового типа, заключены в прямоугольник, виды, содержащие Т-ДНК Ri плазмиды неизвестного типа опинов, подчеркнуты)
Актов трансформации в эволюции рода Nicotiana было несколько и проходили они независимо.")
")
Слайд 44Скрининг видов на наличие Т-ДНК содержащих последовательностей
Закрепление Т-ДНК в геноме
Встречается ли у других родов?
Какова эволюционная роль вставки?
Разработка подхода на основе Real-time PCR:
использование вырожденных праймеров и TaqMan-зондов

Слайд 45п/кл. Ranunculidae
сем. Berberidaceae (1)
сем. Ranunculaceae (1)
п/кл. Hammamelididae
сем. Betulaceae (4)
сем. Fagaceae (3)
сем.
сем. Buxaceae (1)
сем. Moraceae (1)
п/кл. Caryophyllidae
сем. Amaranthaceae (1)
сем. Portulacaceae (1)
п/кл. Dilleniidae
сем. Salicaceae (4)
сем. Tiliaceae (2)
сем. Brassicaceae (5)
сем. Euphorbiaceae (2)
сем. Malvaceae (1)
п/кл. Rosidae
сем. Grossulariaceae (3)
сем. Rosaceae (23)
сем. Fabaceae (3)
сем. Aceraceae (4)
сем. Hyppocastanoceae (1)
сем. Vitaceae (1)
сем. Celasteraceae (1)
сем. Linaceae (1)
сем. Pedaliaceae (1)
сем. Bignoniaceae (1)
сем. Orobanchaceae (1)
п/кл. Asteridae
сем. Asteraceae (13)
сем. Oleaceae (3)
п/кл. Lamiidae
сем. Convolvulaceae (1)
сем. Cuscutaceae (1)
п/кл. Magnoliidae
Схема родства исследованных видов.
90 видов двудольных растений
(29 семейств)
сем. Ranunculaceae (1)п/кл. Hammamelididaeсем. Betulaceae (4)сем. Fagaceae (3)сем. Ulmaceae (1)сем. Buxaceae (1)сем.")
Слайд 46В 2006 году сотрудниками лаборатории ГКИР был обнаружен новый пример горизонтального
В геноме Linaria vulgaris L. выявлены последовательности, гомологичные Agrobacterium rhizogenes.
Linaria vulgaris L.
Проведен целевой поиск подобных мотивов у других представителей рода Linaria

Слайд 47Выявлены последовательности, гомологичные rolB, rolC, ORF13, ORF14 и mis (ген микимопин-синтазы)
Определена
Детально изучена структура вставки
Определено количество вставок
Охарактеризован район интеграции.
Linaria vulgaris L..
Определена их нуклеотидная последовательностьДетально изучена")
Слайд 48Анализ района инсерции у разных линарий.
У разных видов совпадает район интеграции.
«Видообразующий» характер вставки?!

Слайд 49Обнаружение явления косупрессии у растений
(Alexander R. van der Krol, et
Окраска лепестков петуньи, трансформированной генетическими конструкциями: 35S ::DFR и 35S::CHS
Гены DFR и CHS контролируют биосинтез пигментов:
-ген DFR -кодирует дихлорфлавонол-4-редуктазу
-ген CHS -кодирует халкон-синтазу
Окраска лепестков петуньи,")





