- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Биосинтез белка презентация
Содержание
- 1. Биосинтез белка
- 2. Создана молекулярная машина для сборки пептидов
- 4. Интересно, что Синтез одной молекулы белка длится
- 5. Белки – конечный продукт большинства информационных
- 6. Синтез тысяч различных белков в клетке
- 7. Генетический код Синтез белка отличается от других
- 8. Генетический код Генетический код — это способ
- 9. ГЕОРГИЙ АНТОНОВИЧ ГАМОВ В 1954 году опубликовал
- 10. Роберт Уильям Холли (США) Хар Гобинд
- 11. ГЕНЕТИЧЕСКИЙ КОД 1 2 3
- 12. Трансляция- процесс синтеза белка из аминокислот
- 13. Клетка E. coli имеет 15 000
- 14. Между ними образуется щель, через которую
- 15. Бактериальные тРНК содержат от 73 до
- 16. для выполнения адаптерной функции тРНК имеет:
- 17. Плечо D, содержит дигидроуридин, взаимодействие при
- 19. Этапы трансляции Активация аминокислот Инициация Элонгация Терминация и высвобождение Укладка и посттрансляционный процессинг
- 20. Активация аминокислот Mg+2 зависимые аминоацил-тРНК-синтетазы (АРС-аза или
- 22. Активация аминокислот
- 23. Аминоацилирование тРНК приводит к двум результатам:
- 24. Взаимодействие между аминоацил-тРНК –синтетазами и тРНК называют «вторым генетическим кодом».
- 25. Синтез белка начинается с N-конца и
- 26. Для метионина существует один кодон -
- 27. Поэтому у бактерий есть два типа
- 28. Инициаторная ак у бактерий – формилметионин. Инициаторная ак у эукариот – метионин.
- 29. Активация аминокислот
- 30. Для инициации синтеза полипептида у бактерий
- 31. Инициация Инициирующие комплексы: 30S рибосома соединяется с
- 32. Инициаторный кодон 5ꞌAUG попадает в правильную
- 33. Эта последовательность комплементарно связывается с пиримидин-обогащенной
- 34. Бактериальные рибосомы имеют три сайта связывания аминоацил-тРНК: Аминоацильный (А) Пептидильный (Р) Сайт выхода (Е)
- 35. Сайты А и Р образованы обеими
- 36. К комплексу: 30S субъединица рибосомы, IF-3,
- 37. Далее этот крупный комплекс объединяется с
- 38. Инициация В результате образуется функционально-активная 70S- рибосома, называемая инициаторным комплексом.
- 39. Инициация
- 40. Инициация
- 41. элонгация Необходимо: Инициаторный комплекс Аминоацил-тРНК Три растворимых
- 42. В клетке присоединение каждой ак происходит
- 43. На первой стадии элонгации соответствующая аминоацил-тРНК
- 44. ГТФ гидролизуется, комплекс EF-Tu – ГДФ
- 45. Формирование пептидных связей: Пептидная связь формируется
- 46. α-аминогруппа ак в сайте А выступает
- 47. Транслокация – рибосома передвигается на один
- 48. Деацилированная тРНК смещается из Р сайта
- 50. После транслокации дипептидил-тРНК готова к следующему
- 51. Терминация и высвобождение О терминации сигнализирует один
- 52. Стоп-кодон в А сайте рибосомы, подключаются
- 53. RF-3 участвует в высвобождении субъединицы рибосомы.
- 54. Гидролиз ГТФ под действием EG- G
- 56. Из бактериальных и эукариотических клеток можно
- 58. У бактерий процессы транскрипции и трансляции
- 59. Укладка и посттрансляционный процессинг Образованная полипептидная цепь
- 60. Модификации N-конца и C-конца. В
- 61. N-концевая последовательность – для доставки белка
- 62. Модификации некоторых аминокислот: Гидроксильные группы сер,
- 63. Образование дисульфидных мостиков между остатками цистеина.
- 64. К остаткам глу присоединяются карбоксигруппы. При
- 65. Частичный протеолиз – удаление части пептидной
- 66. Присоединение простетических групп: Гем – при
- 67. Присоединение углеводных остатков к остаткам асн
- 68. Трансформирующая активность онкогена ras исчезает при дефекте изопренилирования белка Ras. Применяется в противоопухолевой терапии.
- 69. Лекарственная регуляция синтеза белка Инактивация факторов
- 70. Лекарственная регуляция Нарушение элонгации: тетрациклины блокируют
- 71. Фолдинг белков Фолдинг – это процесс сворачивания
- 72. Фолдинг белков При нарушении функции шаперонов и
- 73. Регуляция синтеза белка Синтез белка регулируется внешними
- 74. Концентрация белка в клетке определяется сложным
- 75. Транскрипция –первая стадия в сложном и
- 76. Гены тех продуктов, которые необходимы клетке
- 77. Инициацию транскрипции РНК-полимеразой регулируют три типа
- 78. Репрессоры блокируют присоединение РНК-полимеразы к промотору. Активаторы усиливают взаимодействие РНК – полимеразы с промотором.
- 79. Белки-репрессоры связываются с определенными участками ДНК
- 80. Регуляция с участием репрессора, подавляющего транскрипцию,
- 81. Эффектор – это небольшая молекула или
- 82. Положительная регуляция - активаторы связываются с
- 83. Многие бактериальные мРНК полицистронные – содержат
- 84. Кластер генов и промотор, регуляторные последовательности
- 85. Регуляция синтеза белка Принципы экспрессии бактериальных генов
- 86. Регуляция синтеза белка Лактозный оперон (lac) содержит
- 87. Ген галактозидпермеазы (Y)–переносит лактозу внутрь клетки.
- 88. В отсутствии лактозы транскрипция lac-оперона подавлена
- 89. Когда в клетке появляется лактоза происходит
- 90. Регуляция синтеза белка Это типичный пример отрицательной
- 92. В присутствии глюкозы экспрессия генов, необходимых
- 93. В отсутствии глюкозы комплекс СRР-сАМР связывается
- 94. Lac- репрессор – отрицательный регуляторный элемент, реагирующий на лактозу. Оба элемента действуют согласованно.
- 95. Когда Lac- репрессор блокирует транскрипцию, комплекс
- 96. В отсутствии комплекса СRР-сАМР открытый комплекс
- 97. Действие глюкозы на СRР опосредовано сАМР.
- 98. По мере снижения концентрации сАМР (при
- 99. Для сильной индукции lac- оперона необходимо
- 101. Так как большинство генов прокариот находятся
- 102. Молекулярные болезни Серповидно-клеточная анемия Замена в 6-ом
- 103. Серповидно-клеточная анемия
- 104. Молекулярные болезни Талассемии – нарушения синтеза цепей Нв. Нарушен процессинг РНК.
- 105. Спасибо за внимание

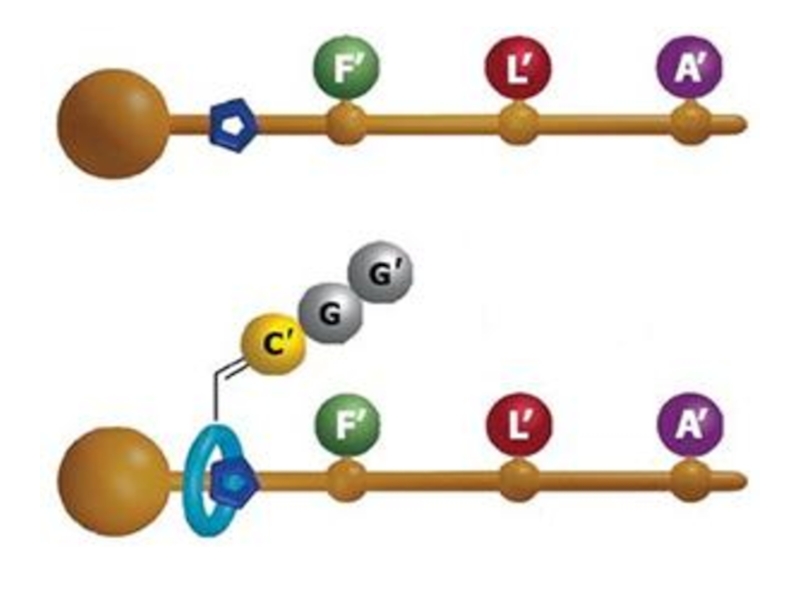
Слайд 2Создана молекулярная машина для сборки пептидов
Исследователи из Манчестера и Эдинбурга
создали примитивный искусственный аналог рибосомы — ротаксановую наномашину, способную синтезировать пептиды заданного состава.

Слайд 4Интересно, что
Синтез одной молекулы белка длится 3-4 минуты.
За одну минуту образуется
от 50 до 60 тыс. пептидных связей.
Половина белков нашего тела
обновляется за 80 дней.
За свою жизнь человек обновляет все свои белки около 200 раз.
Половина белков нашего тела
обновляется за 80 дней.
За свою жизнь человек обновляет все свои белки около 200 раз.

Слайд 5
Белки – конечный продукт большинства информационных метаболических путей.
На синтез белка может
расходоваться до 90% всей энергии клетки.
Полипептид из 100 амк остатков синтезируется в клетке E. coli при 37º С ~5 секунд.
Полипептид из 100 амк остатков синтезируется в клетке E. coli при 37º С ~5 секунд.

Слайд 6
Синтез тысяч различных белков в клетке регулируется таким образом, что их
количество точно соответствует текущему метаболическому состоянию.

Слайд 7Генетический код
Синтез белка отличается от других матричных биосинтезов тем, что между
матрицей и продуктом нет комплементарного соответствия. Поскольку матрица построена из 4 нуклеотидов, а полипептидная цепь — из 20 аминокислот, существует определенный закон шифрования аминокислот в нуклеотидной последовательности матрицы, т.е. генетический код.

Слайд 8Генетический код
Генетический код — это способ записи информации об аминокислотной последовательности
белков с помощью последовательности нуклеотидов в ДНК или РНК.

Слайд 9ГЕОРГИЙ АНТОНОВИЧ ГАМОВ
В 1954 году опубликовал статью,
где первым поднял вопрос
генетического кода,
доказывая, что "при сочетании
4 нуклеотидов тройками
получаются 64 различные
комбинации, чего вполне
достаточно для "записи наследственной
информации"
доказывая, что "при сочетании
4 нуклеотидов тройками
получаются 64 различные
комбинации, чего вполне
достаточно для "записи наследственной
информации"

Слайд 10Роберт Уильям Холли (США)
Хар Гобинд
Корана (США)
Маршалл Уоррен Ниренберг
(США)
За расшифровку генетического кода и его функции в синтезе белков.
 Хар Гобинд Корана (США) Маршалл Уоррен Ниренберг (США) За расшифровку генетического")
Слайд 11
ГЕНЕТИЧЕСКИЙ КОД
1
2
3
ТРИПЛЕТ (КОДОН)
1.ТРИПЛЕТЕН
2.НЕ ПЕРЕК-РЫВАЕТСЯ
АК
ЛЕЙ
3.ОДНОЗНАЧЕН
ГЕН
5.НЕПРЕРЫВЕН
6.УНИВЕРСАЛЕН Б=Г=Р=Ж
1. ОДНА АК КОДИРУЕТСЯ ТРЕМЯ НУКЛЕОТИДАМИ
(ТРИПЛЕТОМ)
2. НУКЛЕОТИД НЕ МОЖЕТ ВХОДИТЬ В СОСТАВ ДВУХ ТРИПЛЕТОВ
3. ТРИПЛЕТ КОДИРУЕТ ТОЛЬКО ОДНУ АК
4. КАЖДАЯ АК ШИФРУЕТСЯ БОЛЕЕ ЧЕМ ОДНИМ КОДОНОМ
5. ВНУТРИ ГЕНА НЕТ ЗНАКОВ ПРЕПИНАНИЯ (СТОП-КОДОНОВ)
6.УНИВЕРСАЛЕН
c к
4.ИЗБЫТОЧЕН (ВЫРОЖДЕН)
1.ТРИПЛЕТЕН2.НЕ ПЕРЕК-РЫВАЕТСЯАКЛЕЙ3.ОДНОЗНАЧЕНГЕН5.НЕПРЕРЫВЕН6.УНИВЕРСАЛЕН Б=Г=Р=Ж1. ОДНА АК КОДИРУЕТСЯ ТРЕМЯ НУКЛЕОТИДАМИ (ТРИПЛЕТОМ)2. НУКЛЕОТИД НЕ МОЖЕТ ВХОДИТЬ")
Слайд 12
Трансляция- процесс синтеза белка из аминокислот на матрице РНК, осуществляемый рибосомальным
комплексом, где ведущую роль играют огромные молекулы РНК.

Слайд 13
Клетка E. coli имеет 15 000 и более рибосом, состоящие из
65% рРНК и 35% белка.
Рибосомы состоят из двух неравных субъединиц: 30S и 50S, содержат очень большие молекулы РНК.
50S состоит из 5S- и 23S-рРНК + 36 белков.
30S состоит из 16S-рРНК + 21 белок.
белки вторичны, покрывают поверхность РНК.
Рибосомы состоят из двух неравных субъединиц: 30S и 50S, содержат очень большие молекулы РНК.
50S состоит из 5S- и 23S-рРНК + 36 белков.
30S состоит из 16S-рРНК + 21 белок.
белки вторичны, покрывают поверхность РНК.

Слайд 14
Между ними образуется щель, через которую при трансляции проходит молекула РНК.
Образование
пептидной связи катализирует рибозим.

Слайд 15
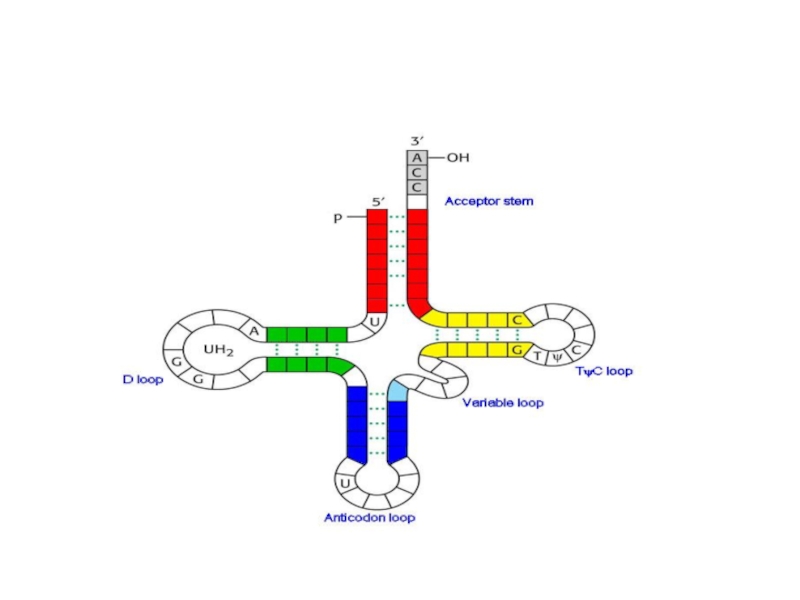
Бактериальные тРНК содержат от 73 до 93 ак.
Каждой амк своя тРНК.
Для
распознавания кодонов всех ак требуется не менее 32 типов тРНК.

Слайд 16
для выполнения адаптерной функции тРНК имеет:
аминокислотное плечо на 3‘конце тРНК.
Антикодоновое плечо
содержит антикодон.
Кодон-триплет нуклеотидов, кодирующих определенную ак.
Кодон-триплет нуклеотидов, кодирующих определенную ак.

Слайд 17
Плечо D, содержит дигидроуридин, взаимодействие при укладке молекул тРНК.
плечо ТΨС, содержащего
риботимидин и псевдоуридин, обеспечивает взаимодействие с большой субъединицей рРНК.

Слайд 19Этапы трансляции
Активация аминокислот
Инициация
Элонгация
Терминация и высвобождение
Укладка и посттрансляционный процессинг

Слайд 20Активация аминокислот
Mg+2 зависимые аминоацил-тРНК-синтетазы (АРС-аза или кодаза)
α-Карбоксил ак атакует
α-фосфат АТФ, образуя 5ꞌ -аминоациладенилат.
Аминоацильная группа переносится на тРНК.
Аминоацильная группа переносится на тРНК.
 α-Карбоксил ак атакует α-фосфат АТФ, образуя 5ꞌ -аминоациладенилат.Аминоацильная")


Слайд 23
Аминоацилирование тРНК приводит к двум результатам:
Активация ак для формирования пептидной связи.
Ак
присоединяется к адаптерной тРНК, что обеспечивает правильное положение ак в растущем полипептиде.

Слайд 24
Взаимодействие между аминоацил-тРНК –синтетазами и тРНК называют «вторым генетическим кодом».

Слайд 25
Синтез белка начинается с N-конца и происходит путем последовательного присоединения ак
к С-концу растущего полипептида.
Инициаторный кодон AUG соответствует N –концевому остатку метионина.
Инициаторный кодон AUG соответствует N –концевому остатку метионина.

Слайд 26
Для метионина существует один кодон - 5ꞌAUG, все организмы имеют для
метионина две тРНК:
Одна тРНК только для инициаторного кодона, другая – для встраивания остатка метионина во внутреннее положение полипептидной цепи.
Одна тРНК только для инициаторного кодона, другая – для встраивания остатка метионина во внутреннее положение полипептидной цепи.




Слайд 30
Для инициации синтеза полипептида у бактерий требуется 30S и 50S –рибосомальные
субъединицы
мРНК
Инициаторная fMet - тРНКfMet
ГТФ, ионы Mg+2
Три белковых фактора инициации:
IF-1,IF-2, IF-3
мРНК
Инициаторная fMet - тРНКfMet
ГТФ, ионы Mg+2
Три белковых фактора инициации:
IF-1,IF-2, IF-3

Слайд 31Инициация
Инициирующие комплексы:
30S рибосома соединяется с IF-1, IF-3.
IF-3 предупреждает преждевременное соединение субъединиц
30S и 50S.
С 30S -субъединицей связывается мРНК.
С 30S -субъединицей связывается мРНК.

Слайд 32
Инициаторный кодон 5ꞌAUG попадает в правильную позицию благодаря последовательности Шайна-Дальгарно в
мРНК.
Эта консенсусная последовательность из 4-9 пуриновых оснований расположена на расстоянии 8-13 п.н. в сторону 5ꞌ-конца от инициаторного кодона.
Эта консенсусная последовательность из 4-9 пуриновых оснований расположена на расстоянии 8-13 п.н. в сторону 5ꞌ-конца от инициаторного кодона.

Слайд 33
Эта последовательность комплементарно связывается с пиримидин-обогащенной последовательностью, находящейся вблизи 3ꞌконца 16S
рРНК 30S –субъединицы рибосомы.

Слайд 34
Бактериальные рибосомы имеют три сайта связывания аминоацил-тРНК:
Аминоацильный (А)
Пептидильный (Р)
Сайт выхода (Е)
Пептидильный (Р)Сайт выхода (Е)")
Слайд 35
Сайты А и Р образованы обеими субъединицами рибосомы 30S и 50S,
сайт Е локализован в 50S субъединице.
Инициаторный 5ꞌ-кодон располагается в пептидильном сайте – здесь связывается fMet - тРНКfMet
Инициаторный 5ꞌ-кодон располагается в пептидильном сайте – здесь связывается fMet - тРНКfMet

Слайд 36
К комплексу: 30S субъединица рибосомы, IF-3, IF-1присоединяется ГТФ и инициаторная fMet
- тРНКfMet
Антикодон этой тРНК связывается с инициаторным кодоном мРНК.
Антикодон этой тРНК связывается с инициаторным кодоном мРНК.

Слайд 37
Далее этот крупный комплекс объединяется с 50S субъединицей рибосомы.
В это же
время ГТФ, связанный с IF-2, гидролизуется до ГДФ и Фн, которые высвобождаются из комплекса.
В этот момент все три фактора инициации отделяются от рибосомального комплекса.
В этот момент все три фактора инициации отделяются от рибосомального комплекса.

Слайд 38Инициация
В результате образуется функционально-активная 70S- рибосома, называемая инициаторным комплексом.



Слайд 41элонгация
Необходимо:
Инициаторный комплекс
Аминоацил-тРНК
Три растворимых цитоплазматических фактора – факторы элонгации (EF-Tu, EF-Ts, EF-G)
ГТФ
ГТФ")
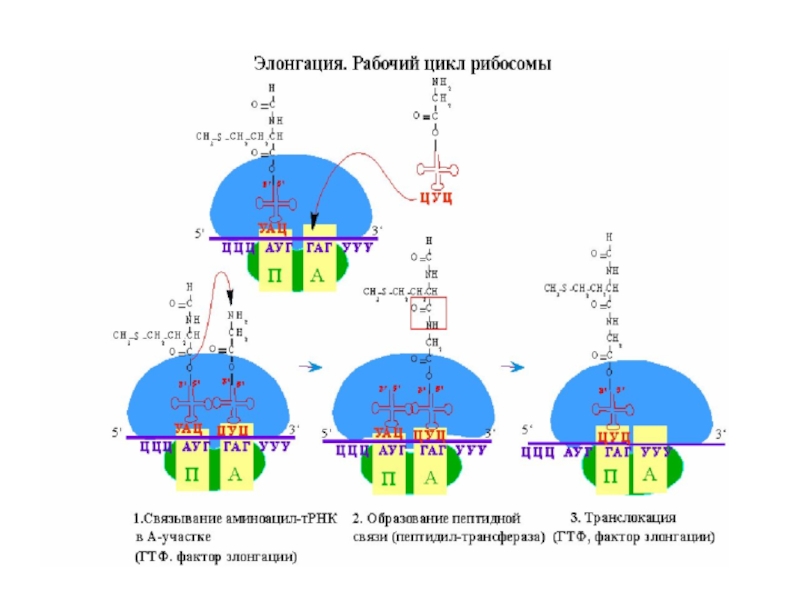
Слайд 42
В клетке присоединение каждой ак происходит в три стадии и эти
стадии повторяются столько раз, сколько ак остатков нужно присоединить.
Элонгация начинается с присоединения второй аминоацил-тРНК.
Элонгация начинается с присоединения второй аминоацил-тРНК.

Слайд 43
На первой стадии элонгации соответствующая аминоацил-тРНК взаимодействует с фактором EF-Tu, связанным
с ГТФ.
Комплекс аминоацил-тРНК - EF-Tu – ГТФ связывается на сайте А 70S инициаторного комплекса.
Комплекс аминоацил-тРНК - EF-Tu – ГТФ связывается на сайте А 70S инициаторного комплекса.

Слайд 44
ГТФ гидролизуется, комплекс EF-Tu – ГДФ высвобождается из 70S рибосомы.
Далее при
участии фактора EF-Ts и ГТФ комплекс EF-Tu – ГТФ регенерирует.

Слайд 45
Формирование пептидных связей:
Пептидная связь формируется между двумя ак, связанными через соответствующие
тРНК с А- и Р-сайтами рибосомы.
Инициаторная N-формилметионильная группа переносится от своей тРНК на аминогруппу второй аминокислоты, находящейся в сайте А.
Инициаторная N-формилметионильная группа переносится от своей тРНК на аминогруппу второй аминокислоты, находящейся в сайте А.

Слайд 46
α-аминогруппа ак в сайте А выступает в роли нуклеофила, вытесняя тРНК
, находящуюся в сайте Р, образуя пептидную связь.
Катализирует реакцию 23S рРНК –пептидилтрансфераза.
В сайте А образуется дипептидил –тРНК , а разгруженная тРНКfMet остается связанной с Р сайтом.
Катализирует реакцию 23S рРНК –пептидилтрансфераза.
В сайте А образуется дипептидил –тРНК , а разгруженная тРНКfMet остается связанной с Р сайтом.

Слайд 47
Транслокация – рибосома передвигается на один кодон по направлению к 3ꞌ
- концу мРНК, при этом антикодон дипептидил-тРНК, который все еще присоединен ко второму кодону мРНК, сдвигается из А сайта в Р сайт.

Слайд 48
Деацилированная тРНК смещается из Р сайта в Е сайт и высвобождается
в цитозоль.
Для транслокации рибосомы вдоль мРНК необходим фактор EF-G и энергия гидролиза ГТФ.
Теперь в А сайте расположен третий кодон мРНК, в Р сайте – второй кодон.
Для транслокации рибосомы вдоль мРНК необходим фактор EF-G и энергия гидролиза ГТФ.
Теперь в А сайте расположен третий кодон мРНК, в Р сайте – второй кодон.

Слайд 50
После транслокации дипептидил-тРНК готова к следующему циклу элонгации и связыванию третьего
аминокислотного остатка.
Таким образом, присоединение одного ак остатка сопровождается гидролизом двух молекул ГТФ до ГДФ и Фн.
Таким образом, присоединение одного ак остатка сопровождается гидролизом двух молекул ГТФ до ГДФ и Фн.

Слайд 51Терминация и высвобождение
О терминации сигнализирует один из трех стоп-кодонов (UAA,UAG,UGA).
Мутации в
антикодоне тРНК, которые позволяют встраивать аминокислоту в ответ на стоп-кодон, губительны для клетки.
.Мутации в антикодоне тРНК, которые позволяют")
Слайд 52
Стоп-кодон в А сайте рибосомы, подключаются три фактора терминации (высвобождения) –
RF-1, RF-2, RF-3.
RF-1 распознает стоп-кодоны UAA,UAG.
RF-2 распознает стоп-кодоны UGA, UAA.
Под действием пептидилтрансферазы растущий полипептид связывается с Н2О.
RF-1 распознает стоп-кодоны UAA,UAG.
RF-2 распознает стоп-кодоны UGA, UAA.
Под действием пептидилтрансферазы растущий полипептид связывается с Н2О.
 – RF-1, RF-2, RF-3.RF-1 распознает")
Слайд 53
RF-3 участвует в высвобождении субъединицы рибосомы.
Высвобождение и распад рибосом на субъединицы
приводит к диссоциации компонентов трансляционного комплекса.
Факторы терминации заменяются на фактор EG-G и фактор рециклизации рибосом (RRF).
Факторы терминации заменяются на фактор EG-G и фактор рециклизации рибосом (RRF).

Слайд 54
Гидролиз ГТФ под действием EG- G приводит к уходу 50S-субъединицы из
комплекса 30S-тРНК-мРНК.
ФакторыEG-G, RRF заменяются на IF-3, который способствует высвобождению тРНК, затем отделяется мРНК.
Затем комплекс 30S субъединица- IF-3 готов инициировать новый раунд синтеза белка.
ФакторыEG-G, RRF заменяются на IF-3, который способствует высвобождению тРНК, затем отделяется мРНК.
Затем комплекс 30S субъединица- IF-3 готов инициировать новый раунд синтеза белка.


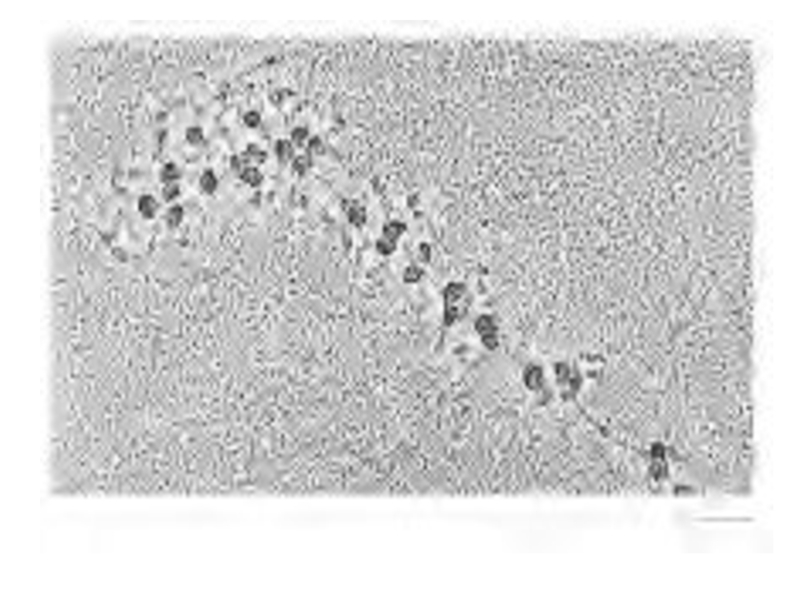
Слайд 56
Из бактериальных и эукариотических клеток можно выделить крупные кластеры из 10-100
рибосом – полисомы, в которых соседние рибосомы соединены между собой тонкими волокнами – молекулами мРНК, с которых происходит трансляция белка одновременно многими рибосомами.
Высокая эффективность процесса.
Высокая эффективность процесса.

Слайд 58
У бактерий процессы транскрипции и трансляции тесно связаны между собой.
Рибосомы начинают
трансляцию (5ꞌ 3ꞌ) еще до завершения транскрипции.
")
Слайд 59Укладка и посттрансляционный процессинг
Образованная полипептидная цепь сворачивается в биологически активную форму.
То
есть линейная белковая молекула превращается в трехмерную структуру.
Но до этого подвергается посттрансляционным модификациям.
У эукариот в ЭР, аппарат Гольджи.
Но до этого подвергается посттрансляционным модификациям.
У эукариот в ЭР, аппарат Гольджи.

Слайд 60
Модификации N-конца и C-конца.
В процессе образования функционального белка формильная группа
(у бактерий), метионин (у эукариот) могут удаляться ферментативным путем.
У эукариот аминогруппа N-конца подвергается ацилированию.
У эукариот аминогруппа N-конца подвергается ацилированию.
, метионин (у")
Слайд 61
N-концевая последовательность – для доставки белка к месту его назначения в
клетке (маркировка)
Удаление сигнальной последовательности специфическими пептидазами.
Удаление сигнальной последовательности специфическими пептидазами.
Удаление сигнальной последовательности")
Слайд 62
Модификации некоторых аминокислот:
Гидроксильные группы сер, тре, тир – фосфорилируются под действием
АТФ, фосфатные группы сообщают полипептидам «-» заряд.
Используется при регуляции активности ферментов, регуляторных белков или для связывания ионов Са2+
Используется при регуляции активности ферментов, регуляторных белков или для связывания ионов Са2+

Слайд 63
Образование дисульфидных мостиков между остатками цистеина.
Дисульфидные мостики защищают нативную конформацию белка
от денатурации во внеклеточной среде.
Метилирование аргинина и лизина в составе гистонов используется для регуляции активности генома
Метилирование аргинина и лизина в составе гистонов используется для регуляции активности генома

Слайд 64
К остаткам глу присоединяются карбоксигруппы.
При участии витамина К происходит γ‑карбоксилирование глутамата
в составе протромбина, проконвертина, фактора Стюарта, Кристмаса.
Позволяет связывать ионы кальция при инициации свертывания крови.
Позволяет связывать ионы кальция при инициации свертывания крови.

Слайд 65
Частичный протеолиз – удаление части пептидной цепи протеолитическими ферментами
Инсулином из проинсулина
Трипсин
из трипсиногена

Слайд 66
Присоединение простетических групп:
Гем – при синтезе гемоглобина, миоглобина, цитохромов, каталазы
Витаминных коферментов
– биотина, ФАД, пиридоксальфосфата и т.п.

Слайд 67
Присоединение углеводных остатков к остаткам асн или сер, тре – гликирование
требуется при синтезе гликопротеинов.
Присоединение изопренильных групп к остатку цис. Помогает заякоривать белок на мембране.
Ras-белки –продукты онкогенов и протонкогенов ras.
Присоединение изопренильных групп к остатку цис. Помогает заякоривать белок на мембране.
Ras-белки –продукты онкогенов и протонкогенов ras.

Слайд 68
Трансформирующая активность онкогена ras исчезает при дефекте изопренилирования белка Ras.
Применяется в
противоопухолевой терапии.

Слайд 69Лекарственная регуляция синтеза белка
Инактивация факторов инициации:
интерферон активирует внутриклеточные протеинкиназы, которые,
в свою очередь, фосфорилируют белковый фактор инициации ИФ‑2 и подавляют его активность.
Нарушение кодон-антикодонового взаимодействия:
стрептомицин присоединяется к малой субъединице и вызывает ошибку считывания первого основания кодона.
Нарушение кодон-антикодонового взаимодействия:
стрептомицин присоединяется к малой субъединице и вызывает ошибку считывания первого основания кодона.

Слайд 70Лекарственная регуляция
Нарушение элонгации:
тетрациклины блокируют А‑сайт рибосомы и лишают ее способности
связываться с аминоацил‑тРНК.
Хлорамфеникол ингибирует пептидилтрансферазу у бактерий; на уэкариот не влияет.
Циклогексимид ингибирует пептидилтрансферазу эукариотических рибосом.
Хлорамфеникол ингибирует пептидилтрансферазу у бактерий; на уэкариот не влияет.
Циклогексимид ингибирует пептидилтрансферазу эукариотических рибосом.

Слайд 71Фолдинг белков
Фолдинг – это процесс сворачивания полипептидной цепи в правильную пространственную
структуру. Для обеспечения фолдинга используется группа вспомогательных белков под названием шапероны (chaperon, франц. – спутник). Они предотвращают взаимодействие новосинтезированных белков друг с другом, изолируют гидрофобные участки белков от цитоплазмы, способствуют переходу вторичной структуры в третичную.

Слайд 72Фолдинг белков
При нарушении функции шаперонов и отсутствии фолдинга в клетке формируются
белковые отложения – развивается амилоидоз. Насчитывают около 15 вариантов амилоидоза.

Слайд 73Регуляция синтеза белка
Синтез белка регулируется внешними и внутренними факторами, которые диктуют
клетке синтез такого количества белка и такого набора белков, которые необходимы для выполнения физиологических функций.

Слайд 74
Концентрация белка в клетке определяется сложным равновесием семи процессов:
Транскрипция
Посттранскрипционная модификация мРНК
Расщепление
мРНК
Трансляция
Посттрансляционная модификация белка
Компартментализация и транспорт белка
Расщепление белка
Трансляция
Посттрансляционная модификация белка
Компартментализация и транспорт белка
Расщепление белка

Слайд 75
Транскрипция –первая стадия в сложном и энергозатратном процессе синтеза белка, поэтому
регуляция концентрации белков как у бактерий, так и эукариот часто осуществляется на уровне транскрипции.

Слайд 76
Гены тех продуктов, которые необходимы клетке постоянно (ферменты основных метаболических путей)
экспрессируются на постоянном уровне в каждой клетке организма (гены домашнего хозяйства).
Регулируемая экспрессия гена – концентрация продуктов этих генов изменяется в ответ на молекулярные сигналы.
Регулируемая экспрессия гена – концентрация продуктов этих генов изменяется в ответ на молекулярные сигналы.
 экспрессируются на постоянном уровне")
Слайд 77
Инициацию транскрипции РНК-полимеразой регулируют три типа белков:
Факторы специфичности изменяют специфичность РНК-полимеразы
по отношению к данному промотору или набору промоторов (σ – субъединица холофермента РНК-полимеразы E.coli).

Слайд 78
Репрессоры блокируют присоединение РНК-полимеразы к промотору.
Активаторы усиливают взаимодействие РНК – полимеразы
с промотором.

Слайд 79
Белки-репрессоры связываются с определенными участками ДНК – операторами.
Оператор часто находится
вблизи промотора.
Репрессор блокирует связывание РНК-полимеразы или ее продвижение вдоль РНК.
Репрессор блокирует связывание РНК-полимеразы или ее продвижение вдоль РНК.

Слайд 80
Регуляция с участием репрессора, подавляющего транскрипцию, называется отрицательной регуляцией.
Связывание репрессора с
ДНК регулируется сигнальной молекулой, эффектором.

Слайд 81
Эффектор – это небольшая молекула или белок, который присоединяется к репрессору
и изменяет его конформацию.
Взаимодействие репрессора с эффектором либо усиливает, либо ослабляет транскрипцию.
Взаимодействие репрессора с эффектором либо усиливает, либо ослабляет транскрипцию.

Слайд 82
Положительная регуляция - активаторы связываются с ДНК и увеличивают активность РНК-полимеразы
на промоторе.
Участки связывания активатора часто примыкают к тем промоторам, с которыми сама (без активаторов) РНК-полимераза не связывается совсем, либо очень слабо.
Участки связывания активатора часто примыкают к тем промоторам, с которыми сама (без активаторов) РНК-полимераза не связывается совсем, либо очень слабо.

Слайд 83
Многие бактериальные мРНК полицистронные – содержат в одном транскрипте несколько генов,
единственный промотор, инициирующий транскрипцию всего кластера.

Слайд 84
Кластер генов и промотор, регуляторные последовательности называются опероном.
Оперон может содержать от
2 до 6 генов, транскрибируемых как единое целое.

Слайд 85Регуляция синтеза белка
Принципы экспрессии бактериальных генов впервые были выявлены при изучении
метаболизма лактозы в клетках E.coli, которая использует этот сахар как единственный источник углерода.
В 1960 г. французские ученые Ф. Жакоб и Ж. Моно лауреаты Нобелевской премии опубликовали статью, где описали регуляцию экспрессии генов, участвующих в метаболизме лактозы у E.coli.
В 1960 г. французские ученые Ф. Жакоб и Ж. Моно лауреаты Нобелевской премии опубликовали статью, где описали регуляцию экспрессии генов, участвующих в метаболизме лактозы у E.coli.

Слайд 86Регуляция синтеза белка
Лактозный оперон (lac) содержит структурные гены, определяющие первичную структуру
синтезируемых белков:
Ген β-галактозидазы (Z)- расщепляет лактозу на галактозу и глюкозу.
Ген β-галактозидазы (Z)- расщепляет лактозу на галактозу и глюкозу.
 содержит структурные гены, определяющие первичную структуру синтезируемых белков:Ген β-галактозидазы (Z)-")
Слайд 87
Ген галактозидпермеазы (Y)–переносит лактозу внутрь клетки.
Ген тиогалактозидтрансацетилазы (A) – модифицирует
токсичные галактозиды для облегчения их удаления из клетки.
Каждому из этих трех генов предшествует участок связывания рибосомы, который направляет трансляцию этого гена независимо от остальных.
Каждому из этих трех генов предшествует участок связывания рибосомы, который направляет трансляцию этого гена независимо от остальных.
–переносит лактозу внутрь клетки. Ген тиогалактозидтрансацетилазы (A) – модифицирует токсичные галактозиды для облегчения")
Слайд 88
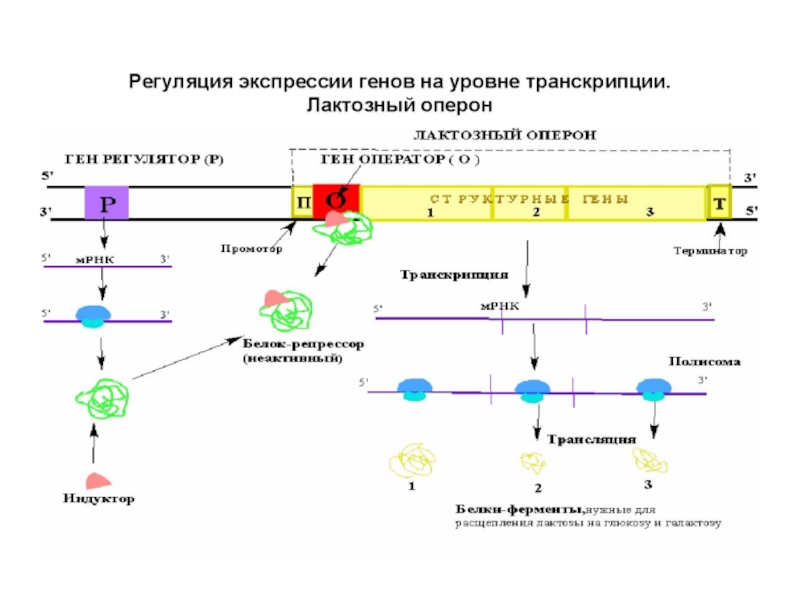
В отсутствии лактозы транскрипция lac-оперона подавлена путем связывания Lac-репрессора.
Lac-репрессор –
это тетрамерный белок, который прочно связывается с оператором.
Lac-репрессор транскрибируется со своего собственного промотора.
Lac-репрессор транскрибируется со своего собственного промотора.

Слайд 89
Когда в клетке появляется лактоза происходит индукция lac-оперона.
Молекула индуктора связывается с
особым участком lac-репрессора, изменяя его конформацию.
Это приводит к отделению репрессора от оператора.
Транскрипция генов lac-оперона.
Это приводит к отделению репрессора от оператора.
Транскрипция генов lac-оперона.

Слайд 90Регуляция синтеза белка
Это типичный пример отрицательной формы регуляции, когда белок-репрессор связывается
с геном-оператором и подавляет транскрипцию.

Слайд 92
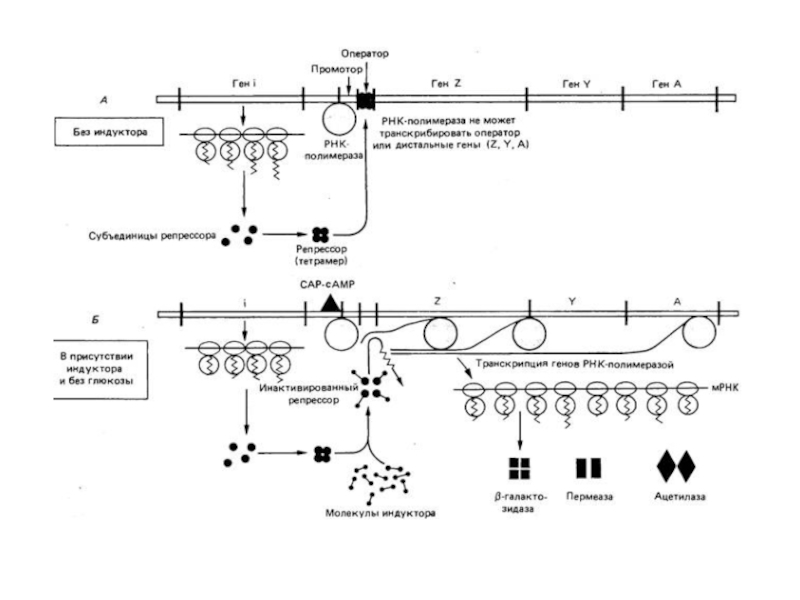
В присутствии глюкозы экспрессия генов, необходимых для катаболизма лактозы, арабинозы ограничивает
регуляторный механизм – катаболитная репрессия.
Влияние глюкозы опосредует сАМР, выступающий как коактиватор.
сАМР-рецепторный белок (СRР) содержит участки связывания ДНК и сАМР.
Влияние глюкозы опосредует сАМР, выступающий как коактиватор.
сАМР-рецепторный белок (СRР) содержит участки связывания ДНК и сАМР.

Слайд 93
В отсутствии глюкозы комплекс СRР-сАМР связывается с ДНК вблизи lac-промотора и
в 50 раз усиливает транскрипцию РНК.
Комплекс СRР-сАМР - положительный регуляторный элемент, реагирующий на концентрацию глюкозы.
Комплекс СRР-сАМР - положительный регуляторный элемент, реагирующий на концентрацию глюкозы.

Слайд 94
Lac- репрессор – отрицательный регуляторный элемент, реагирующий на лактозу.
Оба элемента
действуют согласованно.

Слайд 95
Когда Lac- репрессор блокирует транскрипцию, комплекс СRР-сАМР оказывает незначительное влияние на
Lac-оперон.
Комплекс СRР-сАМР значительно облегчает транскрипцию Lac-оперона при диссоциации репрессора от Lac-оператора.
Комплекс СRР-сАМР значительно облегчает транскрипцию Lac-оперона при диссоциации репрессора от Lac-оператора.

Слайд 96
В отсутствии комплекса СRР-сАМР открытый комплекс РНК-полимеразы и промотора образуется с
трудом.
Белок СRР взаимодействует непосредственно с α – субъединицей РНК-полимеразы.
Белок СRР взаимодействует непосредственно с α – субъединицей РНК-полимеразы.

Слайд 97
Действие глюкозы на СRР опосредовано сАМР.
Наиболее активно СRР связывается с ДНК
при высокой концентрации сАМР, когда концентрация глюкозы низкая.
При высокой концентрации глюкозы синтез сАМР подавляется и стимулируется выход его из клетки.
При высокой концентрации глюкозы синтез сАМР подавляется и стимулируется выход его из клетки.

Слайд 98
По мере снижения концентрации сАМР (при высокой концентрации глюкозы) ослабевает связывание
СRР с ДНК, что снижает экспрессию Lac – оперона.
 ослабевает связывание СRР с ДНК, что")
Слайд 99
Для сильной индукции lac- оперона необходимо присутствие и лактозы (для инактивации
lac-репрессора), и глюкозы в низкой концентрации, что повышает концентрацию сАМР и его связывание с СRР –белком.
, и глюкозы в")
Слайд 101
Так как большинство генов прокариот находятся во «включенном» состоянии, то регуляторные
воздействия направлены на их «выключение».
Для каждого набора генов имеется свой специфический репрессор.
Для каждого набора генов имеется свой специфический репрессор.

Слайд 102Молекулярные болезни
Серповидно-клеточная анемия
Замена в 6-ом положении β- цепи глу на вал,
что приводит к изменению свойств гемоглобина, форма эритроцита меняется ( серп)









