СИСТЕМ И ОРГАНОВ
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Биохимия крови презентация
Содержание
- 1. Биохимия крови
- 2. План лекции: Ферменты плазмы крови. Изоферменты. Конечные
- 3. 1. Ферменты плазмы крови
- 4. Ферменты плазмы крови: 1) Плазмоспецифические; 2) Плазмонеспецифические (внутриклеточные, тканевые, индикаторные).
- 5. Плазмоспецифические ферменты: ЛХАТ(Лецитин: холестерин-ацилтрансфераза)
- 6. Плазмоспецифические ферменты: Липопротеинлипаза
- 7. Плазмоспецифические ферменты: Холинэстераза Лизоцим Факторы свертывания крови
- 8. Диагностическое значение имеет снижение активности фермента
- 9. ПЛАЗМОНЕСПЕЦИФИЧЕСКИЕ ФЕРМЕНТЫ Аспартатаминотрансфераза (АсАТ или АСТ) –
- 10. Внутриклеточное распределение ферментов АлАТ- цитозоль АсАТ- митохондрии,
- 11. Увеличение активности ферментов (гиперферментемия) 1) Усиление синтеза
- 12. Диагностическое значение определения активности внутриклеточных ферментов
- 13. Диагностическое значение определения активности внутриклеточных ферментов
- 14. Понижение активности 1) Снижение числа клеток синтезирующих
- 15. Энзимодиагностика Энзимодиагностика заключается в
- 16. Принципы энзимодиагностики основаны на следующих позициях:
- 17. ряд ферментов имеет преимущественную или абсолютную
- 18. 2. Изоферменты
- 19. Ферменты, катализирующие одну
- 20. По своей структуре
- 21. Изоформы лактатдегидрогеназы (ЛДГ) Фермент
- 22. ЛДГ - олигомерный белок,
- 25. 3. Конечные продукты метаболизма в крови
- 26. Мочевина – конечный продукт
- 27. Мочевина (в сыворотке): 2,50–8,32 ммоль/л
- 28. Образование мочевины (Орнитиновый цикл, Цикл Кребса-Гензелейта)
- 29. Мочевая кислота –
- 30. Мочевая кислота: 179–476 мкмоль/л
- 31. Распад пуриновых нуклеозидов
- 32. Креатинин – содержание
- 33. Креатинин (в сыворотке): Мужчины - 44–150 мкмоль/л Женщины - 44–97 мкмоль/л
- 34. Образование креатинина
- 35. Определение содержания креатина
- 36. Билирубин – относится
- 37. Билирубин: Общий – 3,4–22,2 мкмоль/л Конъюгированный – 0–5,1 мкмоль/л
- 38. 4. Показатели углеводного обмена
- 39. Основной показатель углеводного обмена в крови –
- 41. Для предотвращения чрезмерного
- 42. Виды гипогликемии: Без нарушения функции B-клеток
- 43. снижении катаболизма инсулина в случаях тяжелого
- 44. Виды гипергликемии: Без нарушение функции B-клеток поджелудочной
- 45. Метаболизм глюкозы в эритроцитах
- 46. Метаболизм глюкозы в эритроцитах
- 47. Метаболизм глюкозы в эритроцитах
- 48. 5. Показатели липидного обмена
- 49. Для транспорта липидов кровью
- 50. Трехмерная модель липопротеинов крови
- 51. Основные фракции липопротеинов крови: Хиломикроны (ХМ); Липопротеины
- 53. Все типы липопротеинов
- 54. Холестерин:
- 55. Спасибо за внимание!
Слайд 1БИОХИМИЯ КРОВИ
Лектор:
Чигринский Евгений Александрович
доцент каф. биохимии, кандидат биологических наук
МЕТАБОЛИЧЕСКИЕ ОСОБЕННОСТИ ОТДЕЛЬНЫХ

Слайд 2План лекции:
Ферменты плазмы крови.
Изоферменты.
Конечные продукты метаболизма в крови.
Показатели углеводного обмена.
Показатели липидного
обмена.


Слайд 4Ферменты плазмы крови:
1) Плазмоспецифические;
2) Плазмонеспецифические
(внутриклеточные, тканевые, индикаторные).
 Плазмоспецифические;2) Плазмонеспецифические (внутриклеточные, тканевые, индикаторные).")
")


Слайд 8Диагностическое значение имеет
снижение активности фермента (гипоферментемия)
Признак нарушения функции органа, который
продуцирует фермент
Пусковое звено патологического процесса (дефицит факторов свертывания крови)
Пусковое звено патологического процесса (дефицит факторов свертывания крови)
Признак нарушения функции органа, который продуцирует ферментПусковое звено патологического")
Слайд 9ПЛАЗМОНЕСПЕЦИФИЧЕСКИЕ
ФЕРМЕНТЫ
Аспартатаминотрансфераза (АсАТ или АСТ) – гепатоциты, кардиомиоциты, миоциты.
Аланинаминотрансфераза (АлАТ или АЛТ)
– гепатоциты.
Гамма-глутамилтрансфераза (ГГТ) – клетки канальцев почек, мембраны гепатоцитов.
Креатинфосфокиназа (КФК) – кардиомиоциты, миоциты.
Гамма-глутамилтрансфераза (ГГТ) – клетки канальцев почек, мембраны гепатоцитов.
Креатинфосфокиназа (КФК) – кардиомиоциты, миоциты.
 – гепатоциты, кардиомиоциты, миоциты.Аланинаминотрансфераза (АлАТ или АЛТ) – гепатоциты.Гамма-глутамилтрансфераза (ГГТ)")
Слайд 10Внутриклеточное распределение
ферментов
АлАТ- цитозоль
АсАТ- митохондрии, цитозоль
Аргиназа – ядро,
ГГТ – мембраны,
Лактатдегидрогениза (ЛДГ) –
цитозоль,
Щелочная фосфатаза (ЩФ) – мембраны.
Щелочная фосфатаза (ЩФ) – мембраны.
 – цитозоль,Щелочная фосфатаза (ЩФ)")
Слайд 11Увеличение активности ферментов (гиперферментемия)
1) Усиление синтеза
(ЩФ - при увеличении
давления желчи в желчевыводящих путях).
2) Некроз клеток
(КФК, АсАТ – инфаркт миокарда).
3) Повышение проницаемости мембран
(АсАТ, АлАТ - при вирусном гепатите).
4) Понижение выведения
(ЩФ – при нарушении оттока желчи).
2) Некроз клеток
(КФК, АсАТ – инфаркт миокарда).
3) Повышение проницаемости мембран
(АсАТ, АлАТ - при вирусном гепатите).
4) Понижение выведения
(ЩФ – при нарушении оттока желчи).
1) Усиление синтеза (ЩФ - при увеличении давления желчи в желчевыводящих путях).2)")
Слайд 12
Диагностическое значение определения активности внутриклеточных ферментов в крови
Топическая диагностика (определение места
синтеза)
Сердце – КФК (КК), АсАТ.
Поджелудочная железа – амилаза, липаза.
Предстательная железа – кислая фосфатаза.
Костная ткань – щелочная фосфатаза.
Сердце – КФК (КК), АсАТ.
Поджелудочная железа – амилаза, липаза.
Предстательная железа – кислая фосфатаза.
Костная ткань – щелочная фосфатаза.
Сердце – КФК (КК),")
Слайд 13
Диагностическое значение определения активности внутриклеточных ферментов в крови
Глубина повреждения –
АлАТ, ЛДГ– цитозольные (начальные стадии).
Кислая фосфатаза – лизосомная (глубокое повреждение).
Кислая фосфатаза – лизосомная (глубокое повреждение).
. Кислая")
Слайд 14Понижение активности
1) Снижение числа клеток синтезирующих фермент (цирроз печени проводит к
снижению активности холинэстеразы).
2) Недостаточность синтеза церулоплазмин при болезни Вильсона.
3) Увеличение выведения ферментов (церулоплазмин при нефрозе).
4) Торможение активности трипсина антитрипсином.
2) Недостаточность синтеза церулоплазмин при болезни Вильсона.
3) Увеличение выведения ферментов (церулоплазмин при нефрозе).
4) Торможение активности трипсина антитрипсином.
 Снижение числа клеток синтезирующих фермент (цирроз печени проводит к снижению активности холинэстеразы).2) Недостаточность")
Слайд 15Энзимодиагностика
Энзимодиагностика заключается в постановке диагноза заболевания (или синдрома)
на основе определения активности ферментов в биологических жидкостях человека.
 на основе определения активности ферментов")
Слайд 16
Принципы энзимодиагностики основаны на следующих позициях:
при повреждении клеток в крови или
других биологических жидкостях увеличивается концентрация внутриклеточных ферментов повреждённых клеток;
количество высвобождаемого фермента достаточно для его обнаружения;
активность ферментов в биологических жидкостях, обнаруживаемых при повреждении клеток, стабильна в течение достаточно длительного времени и отличается от нормальных значений;
количество высвобождаемого фермента достаточно для его обнаружения;
активность ферментов в биологических жидкостях, обнаруживаемых при повреждении клеток, стабильна в течение достаточно длительного времени и отличается от нормальных значений;

Слайд 17
ряд ферментов имеет преимущественную или абсолютную локализацию в определённых органах (органоспецифичность);
существуют
различия во внутриклеточной локализации ряда ферментов.
;существуют различия во внутриклеточной локализации")

Слайд 19
Ферменты, катализирующие одну и ту же химическую реакцию,
но отличающиеся по первичной структуре белка, называют изоферментами (изоэнзимами).
Они катализируют один и тот же тип реакции с принципиально одинаковым механизмом, но отличаются друг от друга кинетическими параметрами, условиями активации, особенностями связи апофермента и кофермента.
Они катализируют один и тот же тип реакции с принципиально одинаковым механизмом, но отличаются друг от друга кинетическими параметрами, условиями активации, особенностями связи апофермента и кофермента.

Слайд 20
По своей структуре изоферменты в основном являются олигомерными
белками. Причём та или иная ткань преимущественно синтезирует определённые виды протомеров.
В результате определённой комбинации этих протомеров формируются ферменты с различной структурой - изомерные формы.
Обнаружение определённых изоферментных форм ферментов позволяет использовать их для диагностики заболеваний.
В результате определённой комбинации этих протомеров формируются ферменты с различной структурой - изомерные формы.
Обнаружение определённых изоферментных форм ферментов позволяет использовать их для диагностики заболеваний.

Слайд 21Изоформы лактатдегидрогеназы (ЛДГ)
Фермент ЛДГ катализирует обратимую реакцию окисления
лактата до пирувата:
 Фермент ЛДГ катализирует обратимую реакцию окисления лактата до пирувата:")
Слайд 22
ЛДГ - олигомерный белок, состоящий из 4 субъединиц 2
типов:
М (от англ., muscle - мышца)
Н (от англ., heart - сердце).
Комбинация этих субъединиц лежит в основе формирования 5 изоформ ЛДГ.
ЛДГ1 и ЛДГ2 наиболее активны в сердечной мышце и почках.
ЛДГ4 и ЛДГ5 - в скелетных мышцах и печени. В остальных тканях имеются различные формы этого фермента.
М (от англ., muscle - мышца)
Н (от англ., heart - сердце).
Комбинация этих субъединиц лежит в основе формирования 5 изоформ ЛДГ.
ЛДГ1 и ЛДГ2 наиболее активны в сердечной мышце и почках.
ЛДГ4 и ЛДГ5 - в скелетных мышцах и печени. В остальных тканях имеются различные формы этого фермента.
Н")



Слайд 26
Мочевина – конечный продукт белкового обмена. Повышение содержания мочевины
в крови наблюдается при заболеваниях почек, когда нарушена их выделительная функция, при дегидратации организма и при значительном белковом питании. Снижение содержания мочевины в крови имеет место при заболеваниях печени, когда нарушена ее мочевинообразующая функция.

: 2,50–8,32 ммоль/л")
")
Слайд 29
Мочевая кислота – конечный продукт обмена пуринов. Повышение
содержания – гиперурикемия.
Гиперурикемия может быть: первичная – возникает вследствие нарушения обмена собственно пуринов и вторичная – является сопутствующей основного заболевания или является следствием введения фармпрепаратов, а также характерна для подагры.
Гиперурикемия может быть: первичная – возникает вследствие нарушения обмена собственно пуринов и вторичная – является сопутствующей основного заболевания или является следствием введения фармпрепаратов, а также характерна для подагры.



Слайд 32
Креатинин – содержание в крови небольшое и увеличение
наблюдается при заболевании мышечной ткани и особенно вследствие дистрофических процессов.

: Мужчины - 44–150 мкмоль/л Женщины - 44–97 мкмоль/л")

Слайд 35
Определение содержания креатина и креатинина в крови и
моче используется для характеристики интенсивности работы мышц в спортивной медицине и при некоторых патологических состояниях.
Определение активности фермента креатинкиназы и его изоферментных форм в крови используется в медицине для диагностики таких заболеваний, как инфаркт миокарда, миопатии, мышечные дистрофии и др.
Определение активности фермента креатинкиназы и его изоферментных форм в крови используется в медицине для диагностики таких заболеваний, как инфаркт миокарда, миопатии, мышечные дистрофии и др.

Слайд 36
Билирубин – относится к числу желчных пигментов, образуется
при распаде гемоглобина в клетках РЭС (Ретикуло-эндотелиальная система).



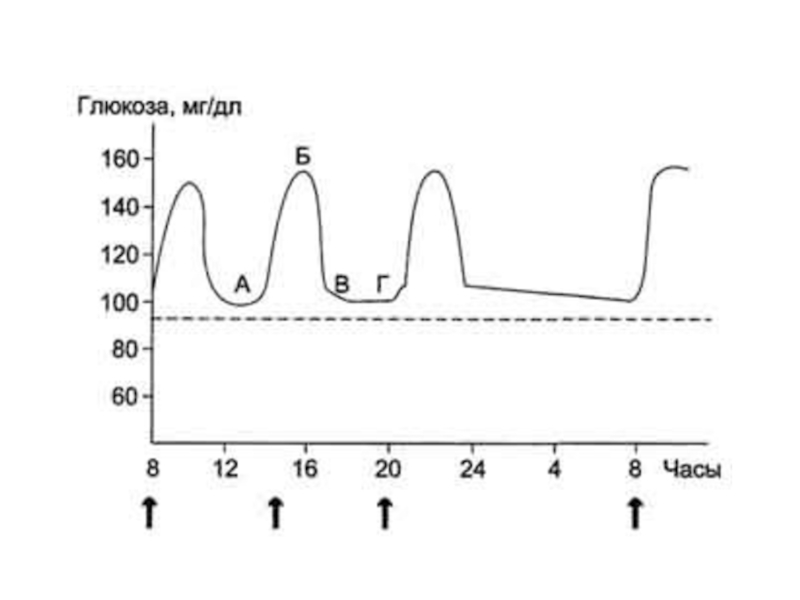
Слайд 39Основной показатель углеводного обмена в крови – концентрация глюкозы
Концентрация глюкозы в артериальной крови в течение суток поддерживается на постоянном уровне 3,3–5,5 ммоль/л.
После приёма углеводной пищи уровень глюкозы возрастает в течение примерно 1 ч до ∼8 ммоль/л, (алиментарная гипергликемия), а затем возвращается к нормальному уровню (примерно через 2 ч).
После приёма углеводной пищи уровень глюкозы возрастает в течение примерно 1 ч до ∼8 ммоль/л, (алиментарная гипергликемия), а затем возвращается к нормальному уровню (примерно через 2 ч).

Слайд 41
Для предотвращения чрезмерного повышения концентрации глюкозы в крови
при пищеварении основное значение имеет потребление глюкозы печенью и мышцами, в меньшей мере - жировой тканью.
Следует напомнить, что более половины всей глюкозы (60%), поступающей из кишечника в воротную вену, поглощается печенью.
Следует напомнить, что более половины всей глюкозы (60%), поступающей из кишечника в воротную вену, поглощается печенью.

Слайд 42Виды гипогликемии:
Без нарушения функции B-клеток
поджелудочной железы
Гипогликемия, не связанная с изменением
эндогенной продукции инсулина наблюдается при:
снижении уровня контринсулярных гормонов (гипокортицизм, гипотиреоз);
передозировке экзогенного инсулина или пероральных гипогликемиических препаратов;
снижении уровня контринсулярных гормонов (гипокортицизм, гипотиреоз);
передозировке экзогенного инсулина или пероральных гипогликемиических препаратов;

Слайд 43
снижении катаболизма инсулина в случаях тяжелого поражения паренхимы печени (гепатит, цирроз,
отравления);
нарушении реабсорбции и потере глюкозы с мочой (снижение почечного порога для глюкозы при почечном диабете);
гликогеновых болезнях (гликогенозы и агликогенозы);
низкоуглеводной диете и голодании.
С нарушением функции B-клеток поджелудочной железы
Наблюдается при повышенной эндогенной продукции инсулина (синдром гиперинсулинизма).
нарушении реабсорбции и потере глюкозы с мочой (снижение почечного порога для глюкозы при почечном диабете);
гликогеновых болезнях (гликогенозы и агликогенозы);
низкоуглеводной диете и голодании.
С нарушением функции B-клеток поджелудочной железы
Наблюдается при повышенной эндогенной продукции инсулина (синдром гиперинсулинизма).
;нарушении реабсорбции и потере")
Слайд 44Виды гипергликемии:
Без нарушение функции B-клеток поджелудочной железы
в результате повышения содержания в
крови гормонов контринсулярного действия.
при сниженной чувствительности тканей к инсулину (сахарный диабет 2 типа).
С нарушением функции B-клеток поджелудочной железы
Обусловлена снижением продукции инсулина: сахарный диабет 1 типа.
при сниженной чувствительности тканей к инсулину (сахарный диабет 2 типа).
С нарушением функции B-клеток поджелудочной железы
Обусловлена снижением продукции инсулина: сахарный диабет 1 типа.

Слайд 45Метаболизм глюкозы в эритроцитах
Эритроциты лишены митохондрий, поэтому в
качестве энергетического материала они могут использовать только глюкозу.
В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов.
Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% - в пентозофосфатном пути.
В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов.
Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% - в пентозофосфатном пути.

Слайд 46Метаболизм глюкозы в эритроцитах
Конечный продукт анаэробного гликолиза лактат
выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа+, К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях.

Слайд 47Метаболизм глюкозы в эритроцитах
Важная особенность анаэробного гликолиза в
эритроцитах по сравнению с другими клетками - присутствие в них фермента бисфосфоглицератмутазы.
Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата.
Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином.
Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата.
Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином.


Слайд 49 Для транспорта липидов кровью в организме образуются комплексы
липидов с белками - липопротеины.


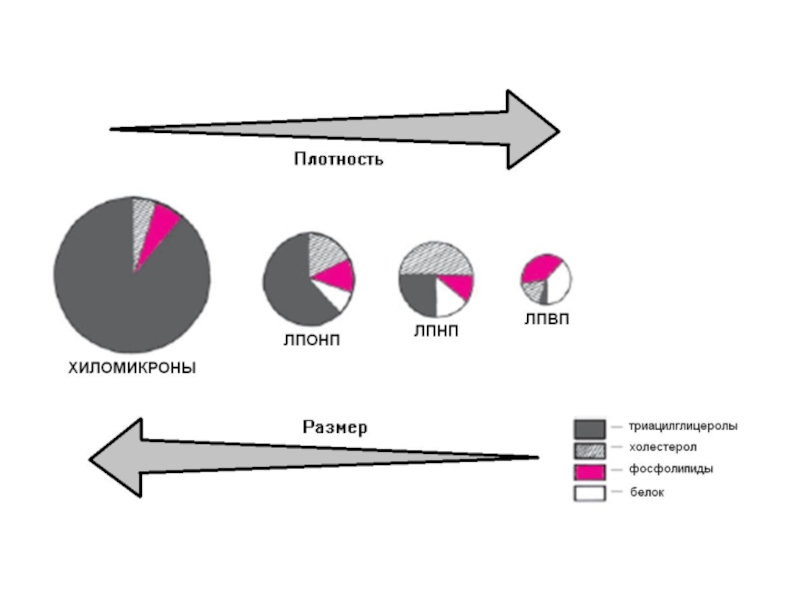
Слайд 51Основные фракции липопротеинов крови:
Хиломикроны (ХМ);
Липопротеины очень низкой плотности (ЛПОНП);
Липопротеины промежуточной плотности
(ЛППП);
Липопротеины низкой плотности (ЛПНП);
Липопротеины высокой плотности (ЛПВП).
Липопротеины низкой плотности (ЛПНП);
Липопротеины высокой плотности (ЛПВП).
;Липопротеины очень низкой плотности (ЛПОНП);Липопротеины промежуточной плотности (ЛППП);Липопротеины низкой плотности (ЛПНП);Липопротеины")
Слайд 53
Все типы липопротеинов имеют сходное строение - гидрофобное
ядро и гидрофильный слой на поверхности. Гидрофильный слой образован белками, которые называют апопротеинами, и амфифильными молекулами липидов - фосфолипидами и холестеролом.
Гидрофильные группы этих молекул обращены к водной фазе, а гидрофобные части - к гидрофобному ядру липопротеина, в котором находятся транспортируемые липиды.
Некоторые апопротеины интегральные и не могут быть отделены от липопротеина, а другие могут свободно переноситься от одного типа липопротеина к другому.
Гидрофильные группы этих молекул обращены к водной фазе, а гидрофобные части - к гидрофобному ядру липопротеина, в котором находятся транспортируемые липиды.
Некоторые апопротеины интегральные и не могут быть отделены от липопротеина, а другие могут свободно переноситься от одного типа липопротеина к другому.








