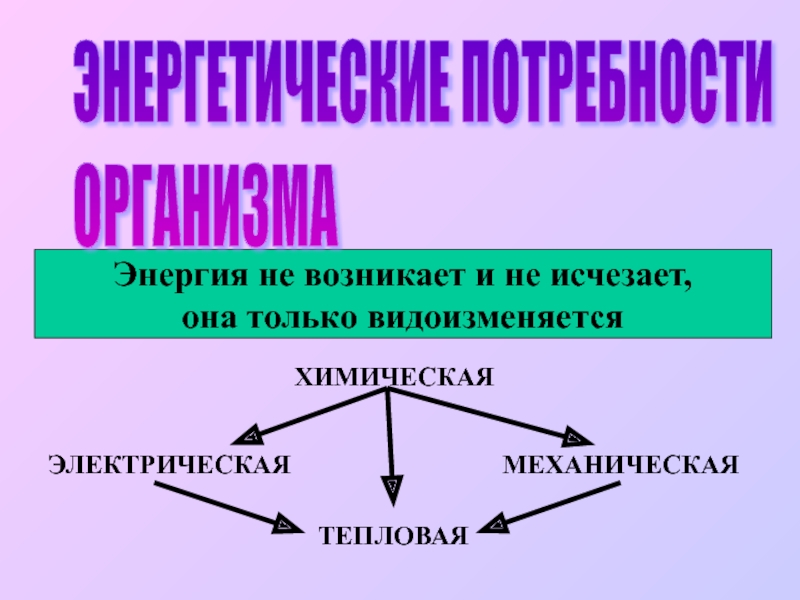
Предшественниками синтеза служат относительно небольшое количество первичных метаболитов.
Синтез четко спланирован и обслуживается набором специальных ферментов, причем некоторые из них весьма специфичны, а некоторые – нет
Для многих групп вторичных метаболитов существует несколько путей их синтеза. Часто этапы синтеза дублированы в разных компартментах клетки (например, пластиды – цитозоль).
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Биохимический моделист-конструктор презентация
Содержание
- 1. Биохимический моделист-конструктор
- 2. Биохимический моделист-конструктор. 1 этап Образуется «скелет» молекулы
- 3. Биохимический моделист-конструктор. 2 этап «Декорирование» скелета:
- 4. Биохимический моделист-конструктор. 2 этап Ацилтрансферазы.
- 5. Образование алкалоидов. Общий взгляд
- 6. Образование пирролидиновых, пиперидиновых и пуриновых алкалоидов
- 7. Образование тропановых алкалоидов
- 8. Образование пирролизидиновых алкалоидов.
- 9. Образование акридоновых алкалоидов
- 10. Образование пуриновых алкалоидов.
- 11. Образование изохинолиновых алкалоидов. Формирование исходной структуры.
- 12. Образование изохинолиновых алкалоидов. Формирование «скелетов»
- 13. Образование изохинолиновых алкалоидов. Протоберберины.
- 14. Синтез изохинолиновых алкалоидов. Морфинаны
- 15. Образование сложных индольных алкалоидов. Исходная структура
- 16. Образование индольных алкалоидов. Формирование «скелетов».
- 17. Ферменты синтеза алкалоидов. H6H, hyoscyamine 6β-hydroxylase;
- 18. Ферменты синтеза алкалоидов. AS, anthranilate synthase;
- 19. Два пути синтеза ИДФ в растениях: «классический» в цитозоле и «альтернативный» в пластидах.
- 21. Синтез изопреноидов. Формирование исходных структур («цепочек»)
- 22. Образование изопреноидов: формирование «скелетов»
- 23. В образовании вторичных метаболитов участвуют сотни ферментов
- 24. Синтез фенольных соединений. Схема синтеза ароматических аминокислот
- 25. Синтез фенольных соединений. Шикиматный путь.
- 26. «Работа» глифосата связана с нарушением транспорта в хлоропласты ERSP-синтазы
- 27. Синтез фенольных соединений. Формирование фенилаланина и тирозина
- 28. Синтез фенольных соединений. Изозимы хоризмат-мутазы
- 29. Синтез фенилпропаноидов
- 30. Синтез фенилпропаноидов.
- 31. Начальные ферменты синтеза фенилпропаноидов Фенилаланин–аммиак-лиаза (ФАЛ, PAL).
- 32. Синтез фенольных соединений. Формирование лигнанов
- 33. Синтез фенольных соединений: пигменты cinnamate-4- hydroxylase (C4H),
- 34. Формирование флавоноидов
- 35. Синтез фенольных соединений. Формирование флавоноидов
- 36. Синтез фенольных соединений. Формирование флавоноидов
- 37. Синтез фенольных соединений. Формирование изофлавоноидов
- 38. Образование беталаинов
- 39. Синтез цианогенных гликозидов
- 40. Образование цианогенных гликозидов
- 41. Образование цианогенных гликозидов
- 42. Образование глюкозинолатов
- 43. «Перекрест» образования глюкозинолатов и цианогенных гликозидов
- 44. Пример метаболической инженерии – вместо глюкозинолатов – амины…
- 45. Как появились ферменты вторичного метаболизма? Модель мембранной
- 46. Основные метаболические пути образования вторичных метаболитов. Часть I.
- 47. Основные метаболические пути образования вторичных метаболитов. Часть II.
- 48. Вторичные метаболизм – дорогое удовольствие…
- 49. Синтез и накопление вторичных метаболитов как правило,
- 50. Внутриклеточная локализация синтеза и хранения вторичных метаболитов
- 51. Места хранения вторичных метаболитов
- 52. Вторичный метаболизм тонко скоординирован в пространстве и
- 53. Аналогично – для хинолизидиновых алкалоидов
- 54. Локализация синтеза тропановых алкалоидов. PMT (путресцин-N-метил трансфераза)
- 55. Локализация синтеза индольных алкалоидов Catharaynthus roseus TDC
- 56. Локализация синтеза монотерпеновых индольных алкалоидов
- 57. Локализация синтеза индольных алкалоидов барвинка
- 58. Метаболизм растительной клетки - причудливое сочетание работы
- 59. Два пути синтеза изопреноидов в растениях: «классический»
- 60. Пластиды и вакуоли – важнейшие игроки в
- 61. Тканевая локализация вторичных метаболитов
- 62. Трихомы и железы листа стевии
- 63. Ультраструктура клеток железки листа стевии
- 64. Летучие вторичные метаболиты львиного зева и петунии
- 65. Возможные пути биосинтеза летучих бензоидов и
- 66. Свет Пути синтеза бензоидов в цветках петунии
- 67. Свет Ночью и днем в цветках петунии
- 68. Что сделали в Японии (группа F.Sato) в
- 69. А затем – уже в дрожжи ввели
- 70. И, наконец… Дополнительная экспрессия в дрожжах генов
- 71. Паклитаксел в дрожжах? Пять генов начальных этапов
- 72. Артемизинин в дрожжах… В Saccharomyces cerevisiae
- 73. Пока единственный пример полного синтеза растительного вторичного
- 74. Зачем нужен вторичный метаболизм растению?
- 75. Стратегия защиты растения от патогенов. Три круга
- 77. ПАТОГЕН системин ферменты терпеноидного метаболизма ФА СК
- 78. Фитоалексины – индуцибельные вторичные метаболиты
- 79. Фитоалексины – индуцибельные вторичные метаболиты
- 80. Фитоалексины – индуцибельные вторичные метаболиты
- 81. Фитоалексины – индуцибельные вторичные метаболиты
- 82. Фитоалексины – индуцибельные вторичные метаболиты
- 83. Фитоалексины – индуцибельные вторичные метаболиты
- 84. Фитоалексины – индуцибельные вторичные метаболиты
- 85. Фитоалексины – индуцибельные вторичные метаболиты
- 86. Фитоалексины – индуцибельные вторичные метаболиты
- 87. Фитоалексины – индуцибельные вторичные метаболиты
- 88. Фитоалексины – индуцибельные вторичные метаболиты
- 89. А как доказать функции не индуцибельных вторичных
- 90. Примеры защитных соединений «первого круга» обороны: алкалоиды
- 91. Примеры защитных соединений «первого круга» обороны: изопреноиды
- 92. Эфирные масла с антимикробиальной активностью
- 93. Эфирные масла с антимикробиальной активностью
- 94. Эфирные масла с антимикробиальной активностью
- 95. Эфирные масла с антимикробиальной активностью против Helicobacter
- 96. Эфирные масла чайного дерева
- 97. Алкалоиды с антимикробиальной активностью против Helicobacter
- 98. Алкалоиды с антимикробиальной активностью против Helicobacter
- 99. Алкалоиды с антимикробиальной активностью против Helicobacter
- 100. Алкалоиды с антимикробиальной активностью против Helicobacter
- 101. Антивирусная активность эфирных масел Plant source
- 102. Примеры защитных соединений «первого круга» обороны: изопреноиды
- 103. Защитное действие смол. Массовая атака короедов сосны
- 104. Примеры защитных соединений «второго круга» обороны: цаногенные гликозиды, глюкозинолаты.
- 105. Примеры защитных соединений «второго круга» обороны:
- 106. Но: постоянное присутствие и биологическая активность не
- 107. Но: постоянное присутствие и биологическая активность не
- 108. Клетки высших растений in vitro - слабовакуолизированные пролиферирующие клетки с небольшим количестовм пластид
- 109. Некоторые закономерности синтеза вторичных метаболитов в культуре
- 110. Клетки высших растений in vitro - гетерогенная
- 111. Вторичный метаболизм в клетках in vitro -
- 112. Синтез фуростаноловых гликозидов в культуре клеток Dioscorea
- 113. Экдистероиды Serratula coronata
- 114. C28 - стероиды
- 115. а - листья интактного растения б
- 116. а - листья интактного растения б
- 117. Женьшень и структура гинзенозидов - тритерпеновых гликозидов даммаранового ряда
- 118. Cструктура семи основных гинзенозидов Гинзенозиды Rg-группы Гинзенозиды Rb-группы
- 119. Содержание гинзенозидов в культуре клеток Panax ginseng
- 120. Содержание гинзенозидов в цикле выращивания суспензионной культуры
- 121. Структура стевиол-гликозидов - тетрациклических дитерпеновых гликозидов стевии
- 122. Содержание стевиол-гликозидов в интактных растениях Stevia
- 123. Содержание стевиол-гликозидов в миксотрофных каллусных культурах стевии
- 124. Ультраструктура клеток каллусных культур стевии
- 125. Вторичный метаболизм – лужок с еще не выщипанной травкой…
Слайд 1Как образуется это разнообразие структур? Общие принципы.

Слайд 2Биохимический моделист-конструктор. 1 этап
Образуется «скелет» молекулы - "базовая" структура, на основе
которой затем формируются многочисленные варианты.
Принципы формирования «скелета» различны для разных групп вторичных метаболитов.
Для изопреноидов – формирование гибких цепочек разной длины - 10, 15, 20, 30, 40 атомов углерода (пренилтрансферазы), а затем сворачивание их в разнообразные циклы (циклазы).
Для алкалоидов – формирование гетероциклов за счет конденсации аминоальдегидов и/или аминокетонов, получаемых из аминов (аминооксидазы), которые в свою очередь формируются из аминокислот (декарбоксилазы)
Принципы формирования «скелета» различны для разных групп вторичных метаболитов.
Для изопреноидов – формирование гибких цепочек разной длины - 10, 15, 20, 30, 40 атомов углерода (пренилтрансферазы), а затем сворачивание их в разнообразные циклы (циклазы).
Для алкалоидов – формирование гетероциклов за счет конденсации аминоальдегидов и/или аминокетонов, получаемых из аминов (аминооксидазы), которые в свою очередь формируются из аминокислот (декарбоксилазы)

Слайд 3Биохимический моделист-конструктор. 2 этап
«Декорирование» скелета: метилирование, ацилирование, десатурация, гидроксилирование, гликозилирование.
«Декорирование» может кардинально менять биологическую активность молекулы.
У арабидопсиса в геноме:
Найдено более 300 (!) генов различных изоформ цитохрома Р450. Трансгенные растения арабидопсиса и табака с двумя чужеродными генами «цианогенных» цитохромов синтезировали цианогенные гликозиды.
Найдены гены более 100 диоксигеназ. Диоксигеназы участвуют в биосинтезе алкалоидов, флавонидов, гиббереллинов, этилена. Для них характерна широкая субстратная специфичность - возможность использовать разные субстраты и образовывать несколько продуктов.
У арабидопсиса в геноме:
Найдено более 300 (!) генов различных изоформ цитохрома Р450. Трансгенные растения арабидопсиса и табака с двумя чужеродными генами «цианогенных» цитохромов синтезировали цианогенные гликозиды.
Найдены гены более 100 диоксигеназ. Диоксигеназы участвуют в биосинтезе алкалоидов, флавонидов, гиббереллинов, этилена. Для них характерна широкая субстратная специфичность - возможность использовать разные субстраты и образовывать несколько продуктов.

Слайд 4Биохимический моделист-конструктор. 2 этап
Ацилтрансферазы. У арабидопсиса – более 50
генов. Ацилтрансферазы вторичного метаболизма у разных видов структурно близки и имеют консервативный гистидин-содержащий мотив. Структурно они также похожи на ацилтрансферазы первичного метаболизма. Гены ацилрансфераз в арабидопсисе собраны в кластеры на 5 хромосоме, тогда как гены диоксигеназ разбросаны по геному
Метилтрансферазы. O-, C-, N-и S- метилтрансферазы, метилируют флавониды, изопреноиды, алкалоиды, полиамины, поликетиды. C-, N-и S- метилтрансферазы эволюционно не связаны, О-метилтрансферазы представляют одну группу и имеют консервативный SАМ-связывающий мотив.
Гликозилтрансферазы. O-, C-, S- гликозилтрансферазы.
Метилтрансферазы. O-, C-, N-и S- метилтрансферазы, метилируют флавониды, изопреноиды, алкалоиды, полиамины, поликетиды. C-, N-и S- метилтрансферазы эволюционно не связаны, О-метилтрансферазы представляют одну группу и имеют консервативный SАМ-связывающий мотив.
Гликозилтрансферазы. O-, C-, S- гликозилтрансферазы.













Слайд 17Ферменты синтеза алкалоидов.
H6H, hyoscyamine 6β-hydroxylase;
ODC, ornithine decarboxylase;
PMT, putrescine-N-methyltransferase;
TR-1, tropinone reductase 1
BBE, berberine bridge enzyme;
CYP80B1, ( S)-Nmethylcoclaurine 3′-hydroxylase;
TYDC, tyrosine decarboxylase.

Слайд 18Ферменты синтеза алкалоидов.
AS, anthranilate synthase;
CPR, cytochrome P450 reductase;
D4H,
desacetoxyvindoline 4-hydroxylase;
DAT, acetyl-CoA:4-O-deacetylvindoline 4-O-acetyltransferase;
DXS, D-1-deoxyxylulose 5-phosphate synthase;
G10H, geraniol 10-hydroxylase;
SGD, strictosidine β-D-glucosidase;
SLS, secologanin synthase;
STR, strictosidine synthase;
TDC, tryptophan decarboxylase;
T16H, tabersonine 16-hydroxylase.
DAT, acetyl-CoA:4-O-deacetylvindoline 4-O-acetyltransferase;
DXS, D-1-deoxyxylulose 5-phosphate synthase;
G10H, geraniol 10-hydroxylase;
SGD, strictosidine β-D-glucosidase;
SLS, secologanin synthase;
STR, strictosidine synthase;
TDC, tryptophan decarboxylase;
T16H, tabersonine 16-hydroxylase.



")

Слайд 23В образовании вторичных метаболитов участвуют сотни ферментов
Синтез изопреноидов:
20 – 30
пренил-трансфераз
100 – 200 циклаз
200 – 300 «декорирующих» ферментов
(оксигеназ, гидроксилаз, метил-, гликозил-, ацил-трансфераз)
100 – 200 циклаз
200 – 300 «декорирующих» ферментов
(оксигеназ, гидроксилаз, метил-, гликозил-, ацил-трансфераз)








Слайд 31Начальные ферменты синтеза фенилпропаноидов
Фенилаланин–аммиак-лиаза (ФАЛ, PAL). Первое сообщение – 1961 год
(юбилей!), из ячменя, МВ ~ 330 kDa, гомотетрамер, Опт. рН ~ 8,8, без кофакторов. Структурно сходна с гистидин-аммиак-лиазой из Pseudomonas putida.
Ряд изоформ. Как минимум две – ФАЛ1 ингибируется кофейной и феруловой кислотами, ФАЛ2 – бензойными кислотами.
Кодируется мультигенным семейством. У Arabidopsis thaliana – 4 гена. Считается растворимым ферментом, однако известны цитоплазматическая и мембрано-связанные изоформы. Предполагается ее ассоциация с гидроксилазой коричной кислоты (С4Н), ассоциированной с ER.
Активируется многими факторами (патогены, УФ).
Для травянистых растений – распространена тирозин-аммиак лиаза (ТАЛ, TAL).
Гидроксилаза коричной кислоты (С4Н, САН). Цитохром-Р450 монооксигеназа. CYP73A1. Известно более 80 цитохром-Р450 семейства CYP73А1, но не все имеют активность С4Н
Мультигенное семейство, Два класса, существенно различающиеся по а-к последовательности, I – для однодольных, II – для двудольных.
Гены клонированы, пробовали их экспрессировать в дрожжах…
Кумарат-СоА-лигаза (4CL). Два класса, I и II. У арабидопсиса – 11 генов, но не все белки проявляют активность. Клонированы, экспрессированы в дрожжах…
Ряд изоформ. Как минимум две – ФАЛ1 ингибируется кофейной и феруловой кислотами, ФАЛ2 – бензойными кислотами.
Кодируется мультигенным семейством. У Arabidopsis thaliana – 4 гена. Считается растворимым ферментом, однако известны цитоплазматическая и мембрано-связанные изоформы. Предполагается ее ассоциация с гидроксилазой коричной кислоты (С4Н), ассоциированной с ER.
Активируется многими факторами (патогены, УФ).
Для травянистых растений – распространена тирозин-аммиак лиаза (ТАЛ, TAL).
Гидроксилаза коричной кислоты (С4Н, САН). Цитохром-Р450 монооксигеназа. CYP73A1. Известно более 80 цитохром-Р450 семейства CYP73А1, но не все имеют активность С4Н
Мультигенное семейство, Два класса, существенно различающиеся по а-к последовательности, I – для однодольных, II – для двудольных.
Гены клонированы, пробовали их экспрессировать в дрожжах…
Кумарат-СоА-лигаза (4CL). Два класса, I и II. У арабидопсиса – 11 генов, но не все белки проявляют активность. Клонированы, экспрессированы в дрожжах…
. Первое сообщение – 1961 год (юбилей!), из ячменя, МВ")

Слайд 33Синтез фенольных соединений: пигменты
cinnamate-4-
hydroxylase (C4H),
chalcone isomerase
(CHI),
chalcone reductase
(CHR),
chalcone
synthase
(CHS),
4-coumaroyl:CoA-
ligase (4CL),
dihydroflavonol
4-reductase (DFR),
7,29-dihydroxy, 49-
methoxyisoflavanol
dehydratase (DMID),
flavanone 3-
hydroxylase (F3H),
flavone synthase
(FSI and FSII),
flavonoid 39
hydroxylase (F39H)
(CHS),
4-coumaroyl:CoA-
ligase (4CL),
dihydroflavonol
4-reductase (DFR),
7,29-dihydroxy, 49-
methoxyisoflavanol
dehydratase (DMID),
flavanone 3-
hydroxylase (F3H),
flavone synthase
(FSI and FSII),
flavonoid 39
hydroxylase (F39H)
, chalcone isomerase(CHI), chalcone reductase (CHR), chalcone synthase (CHS), 4-coumaroyl:CoA-ligase (4CL),")











Слайд 45Как появились ферменты вторичного метаболизма?
Модель мембранной локализации 3-гидрокси-3метилглутарил-КоА-редуктазы ( HMGR).
Белок содержит
вариабельный гидрофобный N-концевой участок (синий), консервативные трансмембранные домены (оранжевые), вариабельные линкерные участки (зеленый и фиолетовый) и консервативный обращенный в цитозоль С-концевой каталитический домен (желтый). Изоформы HMGR, которые определяют синтез вторичных метаболитов после действия элиситоров (сесквитерпеноидных фитоалексинов) отличаются по N-концевому домену и гликозилированы в люмене ЭР.
.Белок содержит вариабельный гидрофобный N-концевой участок")



Слайд 49Синтез и накопление вторичных метаболитов как правило, пространственно разделены
Внутриклеточное разделение:
Синтез:
цитозоль, ЭР, пластиды.
Накопление: вакуоль, периплазматическое пространство
Накопление: вакуоль, периплазматическое пространство
Флуоресценция стероидных гликозидов в клетке
Dioscorea deltoidea in vitro



Слайд 52Вторичный метаболизм тонко скоординирован в пространстве и во времени и на
уровне интактного растения.
Синтез стероидных гликозидов происходит в листьях в водорастворимой (фуростаноловой) форме.
Транспорт осуществляется по флоэме.
Накопление происходит в эпидермальных тканях листьев и стеблей в фуростаноловой форме, в корневищах – в паренхиме в водонерастворимой спиростаноловой форме
Фуростаноловый и спиростаноловые гликозиды отличаются наличием лишь одного остатка глюкозы, но кардинально - по биологической активности.


Слайд 54Локализация синтеза тропановых алкалоидов.
PMT (путресцин-N-метил трансфераза) –
экспрессия в перицикле корня красавки
(A.belladonna)
TR-1 (тропинон редуказа) – локализация в эндодерме и наружном кортексе корня белены (иммунологический анализ)
H6H (гиосциамин -6β-гидроксилаза) – в перицикле корня белены (иммунологический анализ)
Синтез алкалоидов наблюдается только в кончиках молодых корней
(1 – 3 см)
TR-1 (тропинон редуказа) – локализация в эндодерме и наружном кортексе корня белены (иммунологический анализ)
H6H (гиосциамин -6β-гидроксилаза) – в перицикле корня белены (иммунологический анализ)
Синтез алкалоидов наблюдается только в кончиках молодых корней
(1 – 3 см)
 –экспрессия в перицикле корня красавки (A.belladonna)TR-1 (тропинон редуказа) –")
Слайд 55Локализация синтеза индольных алкалоидов Catharaynthus roseus
TDC (триптофан
декарбоксилаза)
STR-1 (стриктозидин
синтаза
локализованы в
клетках верхнего и нижнего
эпидермиса листа.
D4H (дезацетоксивиндолин –гидроксилаза
DAT (деацетилвиндолин
4-О-ацетилтрансфераза) – экспрессируются в идиобластах и млечниках листа.
TDC и STR-1 также экспрессируются в в протодерме корня и кортикальных клетках корня вокруг апикальной меристемы.
Синтез идет только в молодых листьях и корнях.
клетках верхнего и нижнего
эпидермиса листа.
D4H (дезацетоксивиндолин –гидроксилаза
DAT (деацетилвиндолин
4-О-ацетилтрансфераза) – экспрессируются в идиобластах и млечниках листа.
TDC и STR-1 также экспрессируются в в протодерме корня и кортикальных клетках корня вокруг апикальной меристемы.
Синтез идет только в молодых листьях и корнях.
 STR-1 (стриктозидин синтаза локализованы вклетках верхнего и")


Слайд 58Метаболизм растительной клетки - причудливое сочетание работы прокариотческих и эукариотических систем
Синтез
жирных кислот:
согласованная работа многих органелл;
две ацетил-КоА-карбоксилазы:
прокариотческого типа в пластидах,
эукариотического – в цитозоле.
согласованная работа многих органелл;
две ацетил-КоА-карбоксилазы:
прокариотческого типа в пластидах,
эукариотического – в цитозоле.
Синтез флавоноидов:
параллельная работа шикиматного пути в пластидах и цитозоле
Синтез изопреноидов ??

Слайд 59Два пути синтеза изопреноидов в растениях: «классический» (от архебактерий) в цитозоле
и «альтернативный» (от эубактерий) в пластидах.
Моно- сескви- и тритерпеноиды синтезируются по «классическому»
пути в цитозоле клетки
Моно- ди- и тетратерпеноиды - по «альтернативному» в пластидах
Всегда ли?
 в цитозоле и «альтернативный» (от эубактерий)")
Слайд 60Пластиды и вакуоли – важнейшие игроки в метаболическом «пасьянсе»
Пластиды -
«фабрика опасных и энергоемких производств» клетки:
Фотосинтез,
Восстановление нитритов и сульфатов,
Синтез всех жирных кислот,
Синтез аминокислот, пуринов, пиримидинов,
Синтез вторичных метаболитов.
Фотосинтез,
Восстановление нитритов и сульфатов,
Синтез всех жирных кислот,
Синтез аминокислот, пуринов, пиримидинов,
Синтез вторичных метаболитов.
Вакуоли:
Место хранения многих метаболитов, в том числе вторичных (запасающие вакуоли),
Место разрушения метаболитов (литические вакуоли)

Слайд 61Тканевая локализация вторичных метаболитов
А. Сканирующая электронная микрофотография поверхности листа
тимьяна. Круглые структуры - пельтатные железистые волоски (трихомы), в которых синтезируются моно- и сесквитерпены.
B Световая микрофотография железистого волоска мяты, показанного в продольном сечении. C – субкутикулярное пространство; S, секреторные клетки; St - ножка; B, базальная клетка; E, эпидермальная клетка.
C. Световая микрофотография секреторной полости в листе лимона, показанной в поперечном сечении. L, люмен; Sh, клетки футляра; P, клетки паренхимы.
D. Световая микрофотография смоляного хода в древесине сосны Джеффра, показанной в поперечном сечении X, вторичная ксилема.
B Световая микрофотография железистого волоска мяты, показанного в продольном сечении. C – субкутикулярное пространство; S, секреторные клетки; St - ножка; B, базальная клетка; E, эпидермальная клетка.
C. Световая микрофотография секреторной полости в листе лимона, показанной в поперечном сечении. L, люмен; Sh, клетки футляра; P, клетки паренхимы.
D. Световая микрофотография смоляного хода в древесине сосны Джеффра, показанной в поперечном сечении X, вторичная ксилема.



Слайд 64Летучие вторичные метаболиты львиного зева и петунии
myrcene
dimethoxytoluene
Время выхода (min)
Интенсивность
(E)-β-ocimene
linalool
IS
phenylacetaldehyde
Фенилпропаноиды и бензоиды
Изопреноиды
Интенсивность(E)-β-ocimenelinaloolISphenylacetaldehydeФенилпропаноиды и бензоидыИзопреноиды")
Слайд 65Возможные пути биосинтеза летучих бензоидов и
фенилпропаноидов в цветках петунии
2H5
2H5
2H5
2H5
2H5
2H5
2H5
2H5
2H3
2H3

Слайд 66Свет
Пути синтеза бензоидов в цветках петунии ночью и днем
Темнота
Синтез бензойной кислоты
из бензилбензоата является светозависимым процессом.
Orlova et al. (2006) Plant Cell 18

Слайд 67Свет
Ночью и днем в цветках петунии активизируются различные пути биосинтеза бензоидов
Темнота
В
темноте основной вклад в формирование бензоидов вносит
путь синтеза, начинающийся не из фенилаланина…
путь синтеза, начинающийся не из фенилаланина…
Orlova et al. (2006) Plant Cell 18

Слайд 68Что сделали в Японии (группа F.Sato)
в качестве исходного субстрата использовали дофамин,
а в E. coli ввели гены моноамин оксидазы (MAO), норкоклуарин синтазы (NCS), норкоклаурин 6-O-метилтрансферазы (6OMT), коклаурин--N-метилтрансферазы (CNMT) и 3’-гидрокси-N-метилкоклаурин-4’-O-метилтрансферазы (4’OMT). В результате в клетках E. coli был получен синтез коклаурина. Важно, что в экспериментах были использованы гены двух изоформ NCS, но только одна из них показала высокий уровень экспрессии в E. coli.
в качестве исходного субстрата использовали дофамин, а в E. coli")
Слайд 69А затем – уже в дрожжи ввели ген недавно идентифицированной цитохром-Р450-оксидазы
(CYP80G2, коритуберин-синтаза) вместе с генами CNMT и берберин-формирующего энзима (BBE) из Coptis japonica. Трансгенные дрожжи выращивали на среде, содержащей коклаурин, экскретируемый трансгенными клетками E. coli. Показали, что клетки дрожжей содержат магнифлорин и скоулерин. Конечная продуктивность – 55 мг/л (S)-ретикулина, 7,2 мг/л магнифлорина и 8,3 мг/л скоулерина.
 вместе с")
Слайд 70И, наконец…
Дополнительная экспрессия в дрожжах генов трех ферментов (NCS, 6OMT, CNMT)
из Thalictrum flavum, Papaver somniferum и A. thaliana привела к синтезу (S)-скоулерина, (S)-тетрагидроколумбамина и (S)-тетрагидроберберина из (S)-ретикулина. Любопытно, что экспрессия человеческой цитохром-Р450-оксидазы вместе с соответствующей редуктазой привела к формированию из ретикулина салютаридина, показав новую активность введенного фермента…
 из Thalictrum flavum, Papaver")
Слайд 71Паклитаксел в дрожжах?
Пять генов начальных этапов биосинтеза паклитаксела (до образования таксадиен-5α-ацетокси-10β-ола)
были введены в Saccharomyces cerevisiae. Было показано, что ферменты, кодируемые гетерологичными генами, с успехом используют предшественники синтеза изопреноидов дрожжей. Это привело к образованию в клетках дрожжей таксадиена (1 мг/л) и таксадиен-5α-ола (более 25 мкг/л). Однако биосинтез прерывался на стадии гидроксилирования, осуществляющегося цитохром-Р450-оксидазой THY10b…
 были введены в Saccharomyces")
Слайд 72Артемизинин в дрожжах…
В Saccharomyces cerevisiae осуществили экспрессию трех генов - аморфадиен
синтазы, аморфадиен-оксидазы и цитохром Р450-редуктазы - биосинтез от фарнезил-дифосфата до артемизиновой кислоты. Получили артемизиновую кислоту более 100 мг/л.

Слайд 73Пока единственный пример полного синтеза растительного вторичного метаболита в бактерии…
В E.coli
ввели «сборную солянку» генов –
Фенилаланин-аммаик лиазу (PAL) из Rhodotorula rubra
4-Кумарил-KоА-лигазу 4CL из S.coelicor
Халконсинтазу CHS из Glycyrrhiza echinata
Халконизомеразу CHI из Pueraria lobata
Флавоноидов получили мало, но получили.
Однако после оптимизации и всяческих доработок - 700 mg/л флавононов 113 mg/л антоцианинов…
Фенилаланин-аммаик лиазу (PAL) из Rhodotorula rubra
4-Кумарил-KоА-лигазу 4CL из S.coelicor
Халконсинтазу CHS из Glycyrrhiza echinata
Халконизомеразу CHI из Pueraria lobata
Флавоноидов получили мало, но получили.
Однако после оптимизации и всяческих доработок - 700 mg/л флавононов 113 mg/л антоцианинов…

Слайд 74Зачем нужен вторичный метаболизм растению?
Основные точки зрения:
Совсем не нужен
(вторичные метаболиты – «отходы производства»)
Вторичные метаболиты – запасные соединения
Вторичные метаболиты – защитные соединения
Вторичные метаболиты – «биохимические инструменты» взаимодействия растения с окружающей средой
Но как это доказать ?
Вторичные метаболиты – запасные соединения
Вторичные метаболиты – защитные соединения
Вторичные метаболиты – «биохимические инструменты» взаимодействия растения с окружающей средой
Но как это доказать ?
Вторичные")
Слайд 75Стратегия защиты растения от патогенов. Три круга обороны.
Проще всего
доказать защитную роль вторичных метаболитов, участвующих в работе третьего круга обороны: фитоалексины

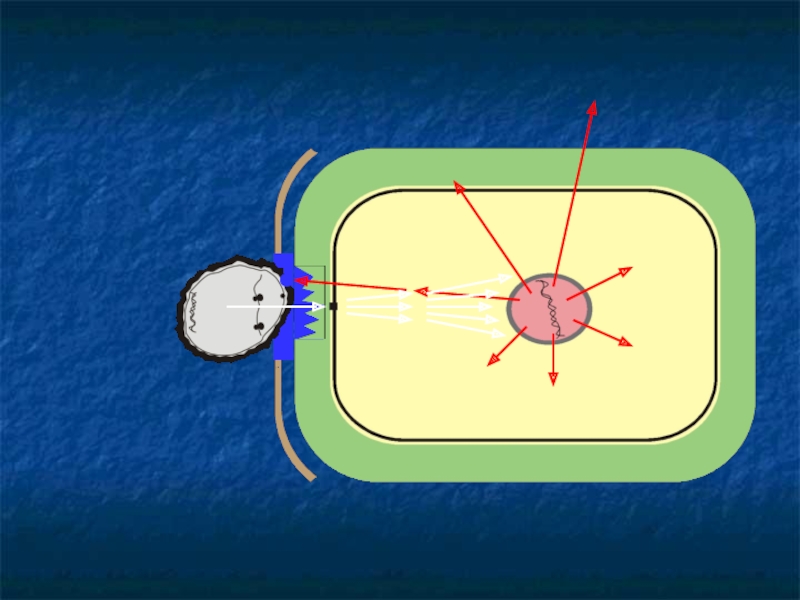
Слайд 77ПАТОГЕН
системин
ферменты терпеноидного метаболизма
ФА
СК
ингибиторы
гликаназ, протеиназ
пролилгидроксилаза
каллозосинтаза
летучие
терпеноиды
цистеиновые
протеиназы
нуклеазы
апоптоз
РИБ
дефензины
хитиназа,
β-1,3 глюаназа
пероксидазы
вакуоль
системный ответ
ферменты фенил-пропаноидного метаболизма
мРНК
К
К
укрепление КС
ОПБ
кал-
лоза
КС












Слайд 89А как доказать функции не индуцибельных вторичных метаболитов?
Авенацин А1 присутствует только
в корнях овса, его нет у пшеницы и ячменя. Мощная защита от патогенного гриба Gaeumannomyces graminis var. tritici, который поражает пшеницу и ячмень, но никогда – овес. У гриба Gaeumannomyces graminis var. avenae есть фермент авенациназа, который удаляет концевые глюкозы и делает молекулу нетоксичной - и этот гриб прекрасно «кушает» овес…
Примеры защитных соединений «первого круга» обороны.


Слайд 91Примеры защитных соединений «первого круга» обороны: изопреноиды
инсектициды: α- и β-пинены,
мирцен, камфен, пиретрин-1
аллелопатические свойства: лимонен, цинеол, карвон, камфора
реппеленты: ментол, непаталактон. α-пинен
против травоядных: 1,8-цинеол
Но – линалоол – привлечение опылителей…
аллелопатические свойства: лимонен, цинеол, карвон, камфора
реппеленты: ментол, непаталактон. α-пинен
против травоядных: 1,8-цинеол
Но – линалоол – привлечение опылителей…
Механическое повреждение – в смоле пихты изменяется содержание пинена, каурена, мирцена.
У устойчивых к абиотическому стрессу растений (загрязнение атмосферы) – много камфена и лимонена, но мало β-пинена, у неустойчивых – наоборот.
После стресса (обработка SO2) – в 5 – 10 раз повышается содержание камфена, α- и β-пинена.










Слайд 101Антивирусная активность эфирных масел
Plant source
Viruses IC50 (%; ppm) References
Herpes simplex virus ( HSV; DNA virus)
Aloysia gratissima HSV-1 65 ppm Garcia et al. (2003)
Artemisia douglasiana HSV-1 83 ppm Garcia et al. (2003)
Citrus limon HSV-1 0.0015% Koch (2005)
Eucalyptus globulus HSV-1 0.009% Schnitzler et al. (2001)
HSV-2 0.008% Schnitzler et al. (2001)
Eupatorium patens HSV-1 125 ppm Garcia et al. (2003)
Houttuynia cordata HSV-1 0.0013% Hayashi et al. (1995)
Hyssopus officinalis HSV-1 0.0001% Koch (2005)
HSV-2 0.0006% Koch (2005)
Illicium verum HSV-1 0.004% Koch (2005)
HSV-.2 0.003% Koch (2005)
Leptospermum HSV-1 0.0001% Reichling et al. (2005
scoparium HSV-2 0.00006% Reichling et al. (2005
Matricaria HSV-1 0.00003% Koch (2005)
recutita HSV-2 0.00015% Koch (2005)
Melaleuca alternifolia HSV-1 0.0009% Schnitzler et al. (2001)
HSV-2 0.0008% Schnitzler et al. (2001)
Mentha piperita HSV-1 0.002% Schuhmacher et al. (2003)
HSV-2 0.0008% Schuhmacher et al. (2003)
Pinus mugo HSV-1 0.0007% Koch (2005)
HSV-2 0.0007% Koch (2005)
Santalum album HSV-1 0.0002% Koch (2005)
HSV-2 0.0005% Koch (2005)
Tessaria absinthioides HSV-1 105 ppm Garcia et al. (2003)
Thymus vulgaris HSV-1 0.001% Koch (2005)
HSV-2 0.0007% Koch (2005)
Zingiber officinale HSV-1 0.0002% Koch (2005)
HSV-2 0.0001% Koch (2005)
Herpes simplex virus ( HSV; DNA virus)
Aloysia gratissima HSV-1 65 ppm Garcia et al. (2003)
Artemisia douglasiana HSV-1 83 ppm Garcia et al. (2003)
Citrus limon HSV-1 0.0015% Koch (2005)
Eucalyptus globulus HSV-1 0.009% Schnitzler et al. (2001)
HSV-2 0.008% Schnitzler et al. (2001)
Eupatorium patens HSV-1 125 ppm Garcia et al. (2003)
Houttuynia cordata HSV-1 0.0013% Hayashi et al. (1995)
Hyssopus officinalis HSV-1 0.0001% Koch (2005)
HSV-2 0.0006% Koch (2005)
Illicium verum HSV-1 0.004% Koch (2005)
HSV-.2 0.003% Koch (2005)
Leptospermum HSV-1 0.0001% Reichling et al. (2005
scoparium HSV-2 0.00006% Reichling et al. (2005
Matricaria HSV-1 0.00003% Koch (2005)
recutita HSV-2 0.00015% Koch (2005)
Melaleuca alternifolia HSV-1 0.0009% Schnitzler et al. (2001)
HSV-2 0.0008% Schnitzler et al. (2001)
Mentha piperita HSV-1 0.002% Schuhmacher et al. (2003)
HSV-2 0.0008% Schuhmacher et al. (2003)
Pinus mugo HSV-1 0.0007% Koch (2005)
HSV-2 0.0007% Koch (2005)
Santalum album HSV-1 0.0002% Koch (2005)
HSV-2 0.0005% Koch (2005)
Tessaria absinthioides HSV-1 105 ppm Garcia et al. (2003)
Thymus vulgaris HSV-1 0.001% Koch (2005)
HSV-2 0.0007% Koch (2005)
Zingiber officinale HSV-1 0.0002% Koch (2005)
HSV-2 0.0001% Koch (2005)
 ReferencesHerpes")
Слайд 102Примеры защитных соединений «первого круга» обороны: изопреноиды
Сесквитерпеноиды:
селинен – инсектицид
Е-β-фарнезен (паслен)
– ферромон тревоги насекомых
Дитерпены:
кауреновая и трахилбановая кислоты (цветки подсолнечника) – гибель личинок бабочек
Паклитаксел (тис) – яд для животных
Тритерпеноиды:
сердечные гликозиды - яд для животных
Экдистероиды – нарушение развития насекомых.
Дитерпены:
кауреновая и трахилбановая кислоты (цветки подсолнечника) – гибель личинок бабочек
Паклитаксел (тис) – яд для животных
Тритерпеноиды:
сердечные гликозиды - яд для животных
Экдистероиды – нарушение развития насекомых.
 – ферромон тревоги насекомыхДитерпены:кауреновая")
Слайд 103Защитное действие смол.
Массовая атака короедов сосны на ствол Pinus contorta. Каждое
белое пятно на стволе - место нападения жуков и выделения смолы из поврежденного ими участка.
Дерево пережило атаку жуков, так как содержание токсичных компонентов эфирного масла, содержащихся в смоле, было достаточно, чтобы убить короедов, которые были затем “удалены” оттоком смолы. После испарения эфирных масел и воздействия воздуха, дитерпеновые смоляные кислоты затвердевают и формируют «пробку», которая «запечатывает» рану.
Дерево пережило атаку жуков, так как содержание токсичных компонентов эфирного масла, содержащихся в смоле, было достаточно, чтобы убить короедов, которые были затем “удалены” оттоком смолы. После испарения эфирных масел и воздействия воздуха, дитерпеновые смоляные кислоты затвердевают и формируют «пробку», которая «запечатывает» рану.


Слайд 105Примеры защитных соединений «второго круга» обороны:
кумарины, фурокумарины
Антимикробное, фунгицидное действие.
Наркотическое
действие – на земляных червей и кроликов
Гипнотическое действие – на мышей.
Кумарины с аминоалкильным заместителями – стимулируют ЦНС.
Псорален, бергаптен – яды для рыб.
Псрален – аллелопатическое действие.
Южные широты – в качестве заместителей – изопреноиды.
Средние широты – кумарины, обогащенные кислородом.
Северные широты – гликозиды, сложные эфиры, метокси- окси-группы.
Псорален – фотосенсибилизатор. 270 и 360 нм.
5 мг на кожу – УФ лампа – через 5 минут сильный ожог.
Граница распространения – между 45 и 55о с.ш.
Севернее – он не встречается. К югу – число видов с псораленом увеличивается.
Гипнотическое действие – на мышей.
Кумарины с аминоалкильным заместителями – стимулируют ЦНС.
Псорален, бергаптен – яды для рыб.
Псрален – аллелопатическое действие.
Южные широты – в качестве заместителей – изопреноиды.
Средние широты – кумарины, обогащенные кислородом.
Северные широты – гликозиды, сложные эфиры, метокси- окси-группы.
Псорален – фотосенсибилизатор. 270 и 360 нм.
5 мг на кожу – УФ лампа – через 5 минут сильный ожог.
Граница распространения – между 45 и 55о с.ш.
Севернее – он не встречается. К югу – число видов с псораленом увеличивается.

Слайд 106Но: постоянное присутствие и биологическая активность не являются достаточным доказательством. Что
делать?
Один из возможных подходов –
использование культур клеток высших растений

Слайд 107Но: постоянное присутствие и биологическая активность не являются достаточным доказательством. Что
делать?
Снятие организменного
контроля развития -
переход к популяционным
механизмам развития системы
Отбор клеток по максимальной и/или устойчивой пролиферации основной принцип развития поуляции
гетерогенность системы по
различным признакам
основа адаптационных возможностей системы
Узкие адаптационые возможности системы
следствие отсутствия длительной эволюции
Большой невостребованный
генетический материал –
клетки in vitro имеют полную
информацию об интактном
растении
Влияние невостребованных
генов на развитие популяции
Судьба невостребованного генетического материала –
возникновение и скорость
мутаций, сомаклональных
вариаций
Включение/выключение невостребованных блоков –
программы морфогенеза,
соматического эмбриогенеза
Основные свойства культуры клеток высших растений -
экспериментально созданной популяции соматических клеток
Один из возможных подходов – использование культур клеток высших растений

Слайд 108Клетки высших растений in vitro - слабовакуолизированные пролиферирующие клетки с небольшим
количестовм пластид

Слайд 109Некоторые закономерности синтеза вторичных метаболитов в культуре клеток высших растений
Избирательная возможность
синтеза метаболитов
не получен синтез морфинанов, димерных индольных алкалоидов, карденолидов.
Различия в синтезе по классам соединений
добиться синтеза фенольных соединений и изопреноидов проще, чем алкалоидов
Различная стабильность синтеза для разных соединений при длительном субкультивирвоании. Стабильный синтез соединений, способствующих пролиферации клеток.
Различные закономерности синтеза в цикле культивирования. Часто синтез активируется при замедлении роста культуры, реже – во время активной пролиферации клеток.
не получен синтез морфинанов, димерных индольных алкалоидов, карденолидов.
Различия в синтезе по классам соединений
добиться синтеза фенольных соединений и изопреноидов проще, чем алкалоидов
Различная стабильность синтеза для разных соединений при длительном субкультивирвоании. Стабильный синтез соединений, способствующих пролиферации клеток.
Различные закономерности синтеза в цикле культивирования. Часто синтез активируется при замедлении роста культуры, реже – во время активной пролиферации клеток.

Слайд 110Клетки высших растений in vitro - гетерогенная популяция соматических клеток
Морфологическая -
клетки табака в суспензии
Биохимическая - флуоресценция клеток диоскореи в культуре
Биохимическая - флуоресценция клеток диоскореи в культуре
Генетическая - различное число хромосом в клетках женьшеня
перестройки хромосом в клетках пшеницы

Слайд 111Вторичный метаболизм в клетках in vitro - синтез изопреноидов
Dioscirea deltoidea
стероидные гликозиды
Panax ginseng, P. japonicus, тритерпновые гликозиды
P.quinquefolius (гинзенозиды)
Serratula coronata, экдистероиды
Ajuga reptans
Stevia rebaudiana дитерпеновые гликозиды
(стевиол-гликозиды)
Polyscias filicifolia тритерпеновые гликозиды
Panax ginseng, P. japonicus, тритерпновые гликозиды
P.quinquefolius (гинзенозиды)
Serratula coronata, экдистероиды
Ajuga reptans
Stevia rebaudiana дитерпеновые гликозиды
(стевиол-гликозиды)
Polyscias filicifolia тритерпеновые гликозиды

Слайд 112Синтез фуростаноловых гликозидов в культуре клеток Dioscorea deltoidea
Протодиосцин
В клетках диоскореи синтезируются
стероидные гликозиды, характерные как для листьев (протодиосцин), так и для корневища (дельтозид) интактного растения. Кроме того появляются 26-S-изомеры, не характерные для интакнтого растения.
S-



Слайд 115а - листья интактного растения
б - длительно культивируемая каллусная культура
в - молодая каллусная культура,
1 - 20-гидроксиэкдизон; 2 - инокостерон; 3 - макистерон А; 5 - экдизон;
ВЭЖХ хроматограммы экдистероидов интактного растения и культур клеток серпухи Serratula coronata
Состав экдистероидов длительно культивируемого каллуса приближается к составу этих соединений в листьях растения

Слайд 116а - листья интактного растения
б - длительно культивируемая каллусная культура
в - молодая каллусная культура,
1 - полиподин В; 2 - 20-гидроксиэкдизон; 3 - 29-норциастерон;
4 - 29-норсенгостерон; 5 - сенгостерон; 6 - аюгалактон; 7 - аюгастерон В;



Слайд 119Содержание гинзенозидов в культуре клеток Panax ginseng в трех последовательных субкультивированиях
на двух разных средах
%
%
Среда ТО
Среда 62

Слайд 120Содержание гинзенозидов в цикле выращивания суспензионной культуры клеток Panax japonicus (repens)
Общее
содержание гинзенозидов
Соотношение Rg/Rb групп
Соотношение Rg/Rb групп
Общее содержание гинзенозидов Соотношение")
Слайд 121Структура стевиол-гликозидов - тетрациклических дитерпеновых гликозидов стевии
Стевиозид =
R1: Glu
R2: Glu- Glu
Ребаудиозид А = R1: Glu
R2: Glu- Glu
|
Glu
Ребаудиозид С = R1: Glu
R2: Glu- Rha
|
Glu
R2: Glu- Glu
Ребаудиозид А = R1: Glu
R2: Glu- Glu
|
Glu
Ребаудиозид С = R1: Glu
R2: Glu- Rha
|
Glu

Слайд 122Содержание стевиол-гликозидов в интактных растениях Stevia rebaudiana и культурах in vitro
(mg/g)
1. Intact plant 1. Callus culture
2. Plant in vitro 2. Suspension culture
3. “Roller” plant 3. Morphogenic callus
4. Shoots from morphogenic callus
 1. Intact plant")

Слайд 124Ультраструктура клеток каллусных культур стевии
Миксотрофная каллусная культура
Гетеротрофная каллусная
культура
Гетеротрофная каллусная
культура