- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Биохимические процессы нервной ткани презентация
Содержание
- 1. Биохимические процессы нервной ткани
- 3. Структура нервных клеток Нервная клетка
- 7. Функции нервной ткани Генерация электрического сигнала
- 8. Химический состав мозга
- 9. На долю головного мозга приходится 2% от
- 10. Верхний ряд - серое вещество в мозге
- 12. 40% сухой массы мозга приходится на
- 13. Ферменты выделенные из ЦНС в кристаллическом
- 14. Образование глутамина – основной путь обезвреживания NH3
- 15. образуют функциональную непрерывность содержат до 50
- 16. Свойства регуляторных пептидов. Дистантные регуляторы. Сложные
- 17. Центральное действие нейропептидов заключается в их
- 18. Опиоидные регуляторные пептиды. α-эндорфин – стимулятор
- 19. Белки миелиновой оболочки Основной белок
- 20. Циклические нуклеотиды участвуют в синаптической передаче
- 21. Нуклеиновые кислоты ДНК не синтезируется
- 22. Углеводы Ткань мозга бедна
- 23. Липиды Находятся в клеточных и субклеточных мембранах
- 24. 25 г холестерина в головном мозге взрослого,
- 25. Энергетический обмен головного мозга
- 26. Запасы гликогена в клетках головного мозга
- 28. Потенциал покоя и потенциал действия
- 29. Потенциал покоя Мембраны, в том чикле плазматические,
- 32. Потенциал действия Возбуждение нервной клетки под действием
- 33. медиаторы нервной системы
- 35. Нейромедиаторы и нейрогормоны Нервные клетки управляют
- 36. Критерии, по которым вещество может быть отнесено
- 38. Химическое строение По химическим свойствам нейромедиаторы
- 39. Ацетилхолин – сложный эфир уксусной кислоты и
- 41. Норадреналин действует на адренэргические рецепторы. Повышение цАМФ
- 42. Дофамин, серотонин, ГАМК. Депрессия связана
- 43. Глицин – медиатор торможения, подобный ГАМК. Функционирует
- 45. Синапсы
- 47. В сетчатке глаза позвоночных содержатся
- 49. Фоторецептор На рисунке схематически изображена одна
- 51. Сигнальный каскад
- 52. В темноте (на схеме слева внизу) высокий
- 53. Спасибо за внимание!

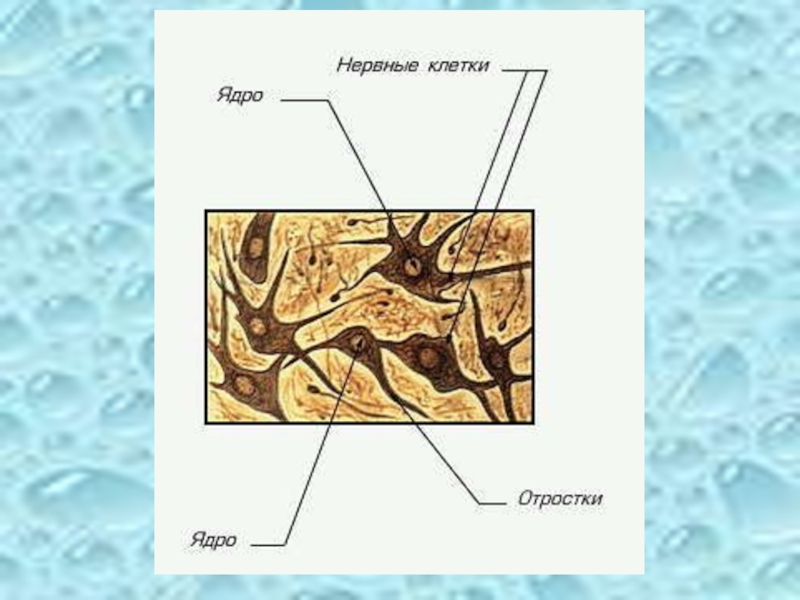
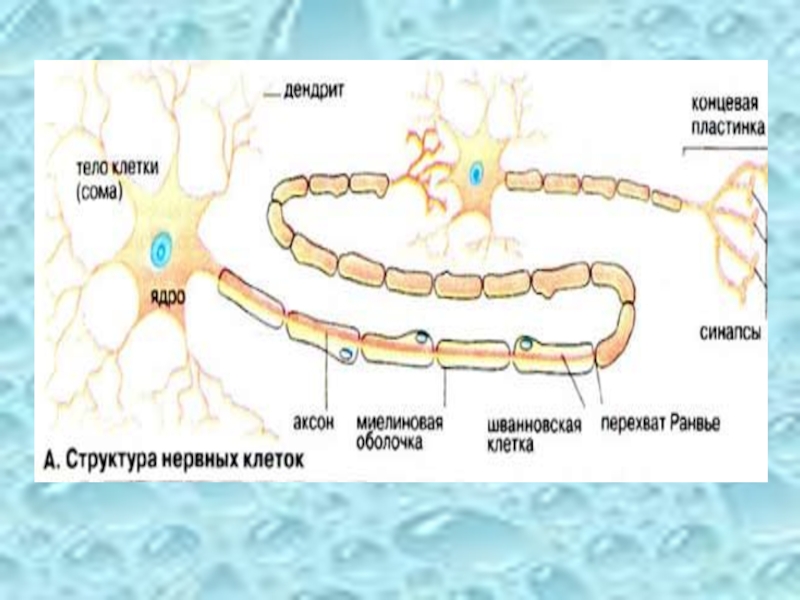
Слайд 3Структура нервных клеток
Нервная клетка состоит из:
тела клетки (сомы)
отростков (аксонов
концевых пластинок
С помощью дендритов нейроны воспринимают, а посредством аксонов передают возбуждение. На периферии аксоны покрыты шванновскими клетками, образующими миелиновую оболочку с высокими изолирующими свойствами.
отростков (аксонов и дендритов) концевых пластинок")
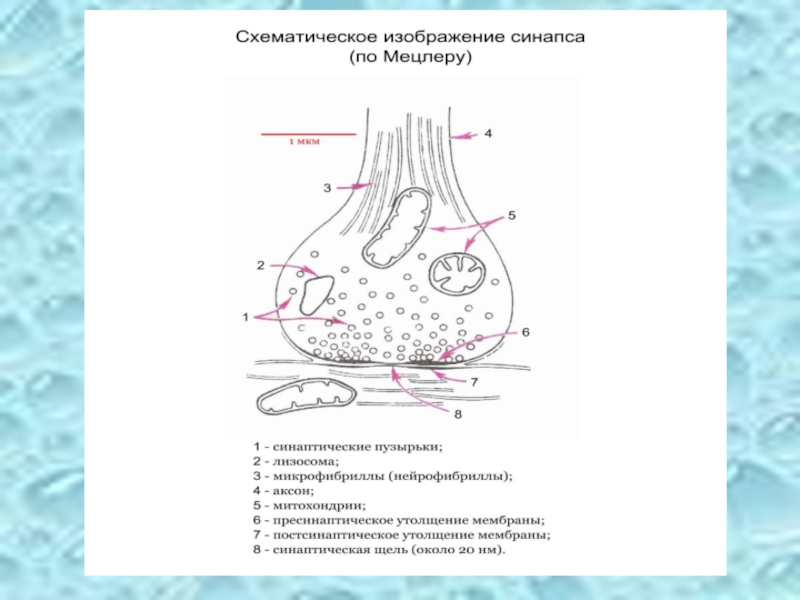
Слайд 7Функции нервной ткани
Генерация электрического сигнала (нервного импульса)
Проведение нервного импульса
Запоминание и хранение
Формирование эмоций и поведения
Мышление
Проведение нервного импульсаЗапоминание и хранение информацииФормирование эмоций и поведенияМышление")

Слайд 9На долю головного мозга приходится 2% от массы тела. 25% составляет
При прекращении доступа O2 мозг может «просуществовать» немногим более 6 минут за счёт резерва лабильных фосфатов. АТФ, креатинфосфат, характеризуются значительным постоянством в головном мозге. Интенсивность обновления фосфорных соединений в головном мозге велика.

Слайд 10Верхний ряд - серое вещество в мозге мужчины и женщины Нижний
Серое вещество головного мозга - тела нейронов.
Белое вещество – аксоны.
В сером веществе 84% H2O, в белом 70%. Белки составляют ½ объёма плотного вещества в сером веществе, в белом 1/3. Липиды составляют больше ½ от сухого остатка в белом веществе и 1/3 в сером.

Слайд 12 40% сухой массы мозга приходится на белки. Более 100 белков
Простые белки
Нейроальбумины (на них 90% от всех белков), нейроглобулины, нейросклеропротеины (в беловом веществе), нейроколлаген, нейроэластины, нейростромины.
Сложные белки
Нуклеопротеины, липопротеины, протеолипиды (липидный компонент преобладает над белковым, сосредоточены в миелине) фосфопротеины, гликопротеины.
В мозговой ткани содержатся в значительном количестве ещё более сложные надмолекулярные образования, такие как липонуклеопротеины, липогликопротеины, липогликонуклеопротеиновые комплексы
Белки мозга

Слайд 13 Ферменты выделенные из ЦНС в кристаллическом виде ацетилхолинэстераза и креатинфосфокиназа.
До 75% АМК мозга представлены глу, асп и их производными ( N-ацетиласпарогиновая, глутамин, глутатион, ГАМК).Много таурина и цистатианина в ткани мозга

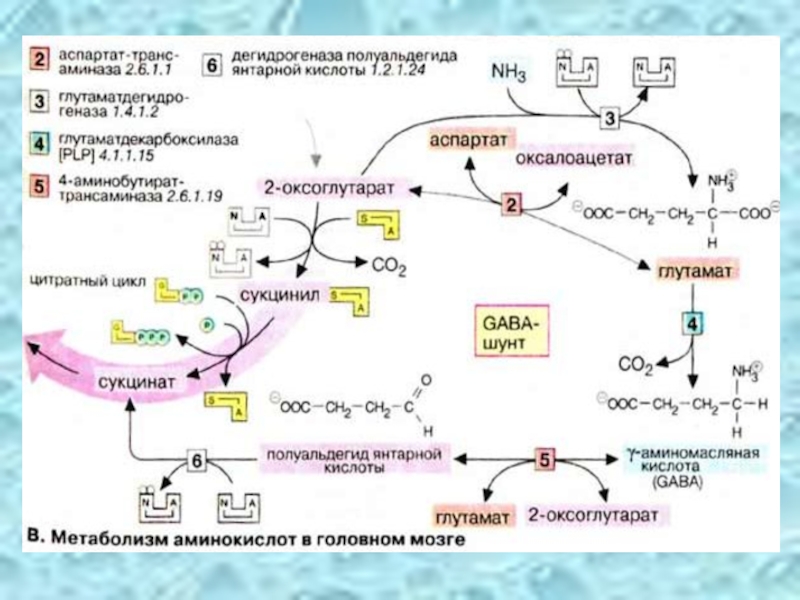
Слайд 14Образование глутамина – основной путь обезвреживания NH3 в ткани мозга.
Белки
Действие глутамата на синаптосомы мозга характеризуется увеличением входа 45Ca, повышением активности NO-синтазы, усилением образования активных форм кислорода, окислением тиолов, ингибированием Na, K – АТФазы и выходом из синаптосом ЛДГ, что может быть причиной повреждающего (нейротоксического) эффекта глутамата.

Слайд 15образуют функциональную непрерывность
содержат до 50 аминокислотных остатков.
переносят информацию в
осуществляют дистантную регуляцию.
Схема образования регуляторных пептидов
из неактивного белка-предшественника про-опиомеланокортина (ПОМК).
Регуляторные пептиды

Слайд 16Свойства регуляторных пептидов.
Дистантные регуляторы.
Сложные функции.
В плазме регуляторных пептидов
Наличие рецепторы для регуляторных пептидов.
Ничтожная концентрация регуляторных пептидов вызывает мощный эффект 10-12 – 10-14 Брадикинин 10-16 М. Эндотелин приводит к спазму коронарных сосудов.
Наличие высокой и специфической активности у продуктов неполного протеолиза регуляторных пептидов.
Сроки полураспада регуляторных пептидов от минут до часов.
Регуляторные пептиды образуются из пептидов-предшественников протеазами.
Любой пептид биологически активен и вмешивается в регуляцию

Слайд 17Центральное действие нейропептидов
заключается в их способности выступать
в качестве как
нейромодуляторов, изменяющих функциональнуюактивность нейрона.
Нейропептиды карнозин и гомокарнозин выполняют в ткани мозга функцию гидрофильных антиоксидантов. Карнозин поддерживает гомеостаз организма при экстремальных воздействиях.
Гипоталамические либерины и статины.
Соматолиберин индуцирует СТГ.
Люлиберин активирует половое
поведение и секрецию
гонадотропонина и ЛГ.

Слайд 18Опиоидные регуляторные пептиды.
α-эндорфин – стимулятор эмоционального поведения и двигательной активности. Опиоидная
β-эндорфин – опиоид, анальгезия, ретроградная амнезия.
γ-эндорфин – нейролептик, опиоидные свойства мало выражены.
К нейропептидам относятся: вазопрессин, окситоцин, панкреатические пептиды,(глюкагон), нейротензины, кинины, ангиотензины, кальцитонин.
Энкефалины – эндогенные антистрессовые биорегуляторы обладают отчётливым противошоковым эффектом.

Слайд 19Белки миелиновой оболочки
Основной белок (30% от общего белка миелина). Если
Протеолипидная белковая фракция.
Кислый протеолипид. В миелине обнаруживается цАМФ-зависимая киназа, которая фосфорилирует белок миелина, и фосфодиэстераза, гидролизующая цАМФ. Период полужизни миелина – 1 месяц.
1 - ядро; 2 – тело клетки;
3 - дендрит; 4 – перехваты Ранвье;
5 - аксон; 6 – миелиновая оболочка; 7 – нерв с сотнями аксонов;
. Если блокировать этот белок то")
Слайд 20Циклические нуклеотиды
участвуют в синаптической передаче нервного импульса. Много цАМФ и
Минеральные вещества
Фосфора в белом веществе больше, чем в сером. В ткани мозга дефицит анионов. Он покрывается за счёт липидов. Участие липидов в ионном балансе одна из их функций в деятельности головного мозга.

Слайд 21Нуклеиновые кислоты
ДНК не синтезируется
Содержание и скорость синтеза РНК большая
Пиримидиновые нуклеотиды
Пуриновые нуклеотиды синтезируются в нервной ткани
Циклические нуклеотиды (цАМФ, цГМФ) содержатся в нервной ткани в большом количестве

Слайд 22Углеводы
Ткань мозга бедна углеводами (гликоген, глюкоза) 100-120 гр. глюкозы
Кетоновые тела используются мозгом лишь при голодании и длительной работе. Метаболизм мозга за счёт резерва углеводов долго продержаться не может, так как содержание гликогена здесь невелико (0,1%). С этим связано развитие комы при избыточном введении инсулина. В ткани мозга активны ГК, ФФК.
 100-120 гр. глюкозы в сутки используется в ткани")
Слайд 23Липиды
Находятся в клеточных и субклеточных мембранах нейронов и в миелиновых оболочках.
В мозговой ткани обнаружены ганглиозиды и немного нейрального жира. В миелиновой оболочке 70-80% липидов и 20-30% белков. Миелин – изолятор, обеспечивает в 6-ть раз более быстрый перенос нервного импульса, чем в немиелинизированных волокнах. В миелиновой оболочке отношение холестерин : фосфоглицериды : галактолипиды составляет 4 : 3 : 2. Основной фосфоглицерид - фосфатидилэтаноламины. Основной галактолипид – цереброзиды. Содержание сфингомиелинов невелико в головном мозге и выше в миелине периферических нервов. Скорость обновления липидов низкое, медленно протекает механизм холистерина, цереброзидов, фосфатидилэтаноламинов, сфингомиелинов.

Слайд 2425 г холестерина в головном мозге взрослого, 2 г у новорождённого.

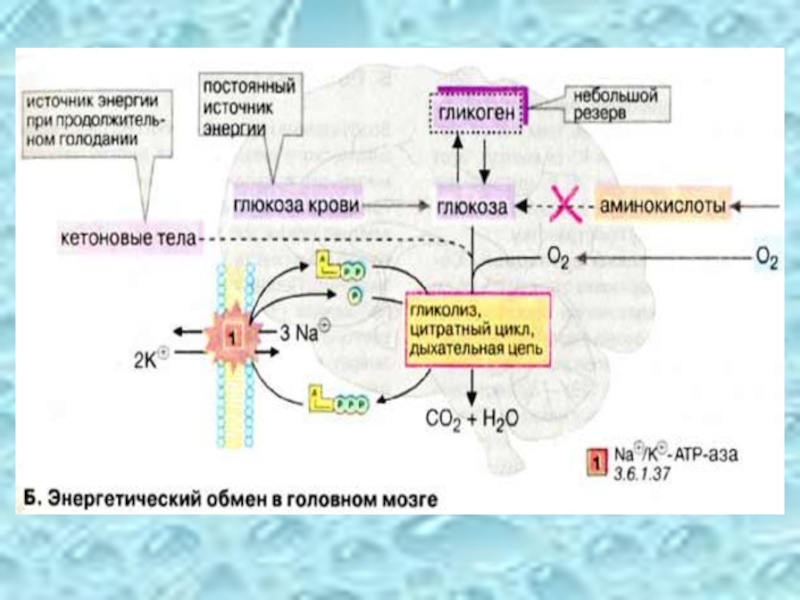
Слайд 25Энергетический обмен головного мозга
Головной мозг хорошо снабжается кровью и имеет интенсивный
В клетках головного мозга практически единственным источником энергии, который должен поступать постоянно, является глюкоза. Только при продолжительном голодании клетки начинают использовать дополнительный источник энергии — кетоновые тела.

Слайд 26 Запасы гликогена в клетках головного мозга незначительны.
Жирные кислоты, которые
Аминокислоты не могут служить источником энергии для синтеза АТФ (АТР), поскольку в нейронах отсутствует глюконеогенез.
Зависимость головного мозга от глюкозы означает, что резкое падение уровня глюкозы в крови, например, в случае передозировки инсулина у диабетиков, может стать опасным для жизни.
глиальные клетки способны реагировать на обмен сигналами между нейронами.


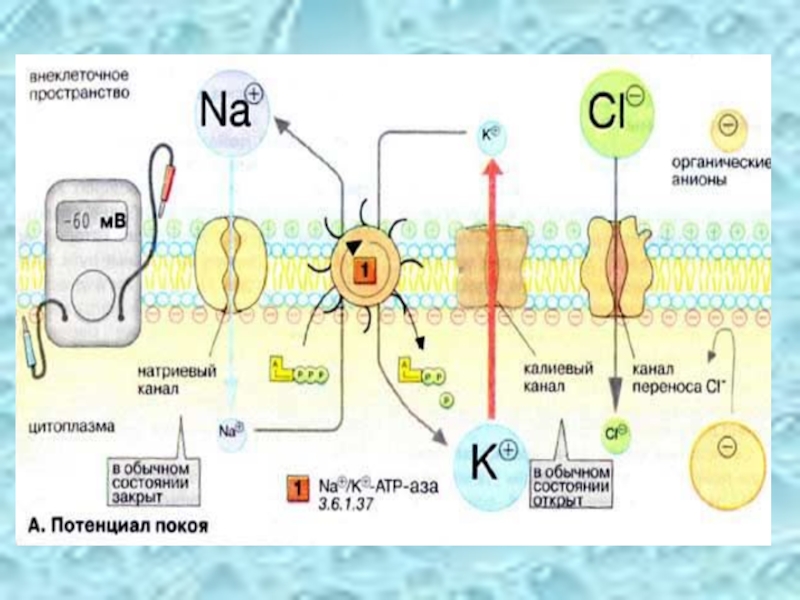
Слайд 29Потенциал покоя
Мембраны, в том чикле плазматические, в принципе непроницаемы для заряженных

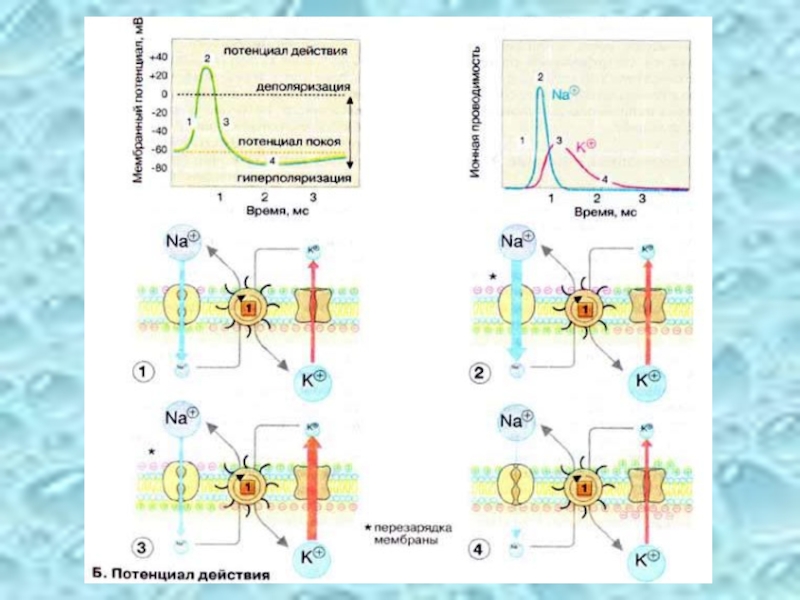
Слайд 32Потенциал действия
Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса)
За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоих ионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
 приводит к возникновению потенциала")

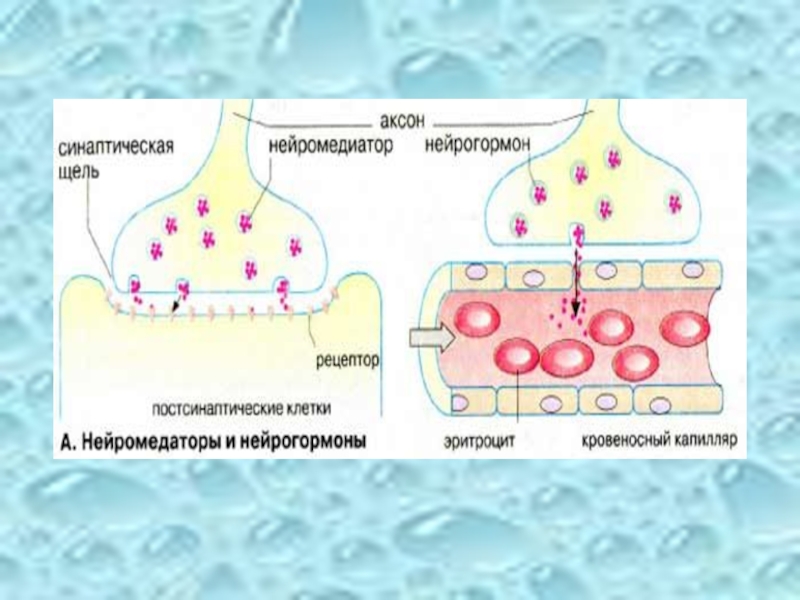
Слайд 35Нейромедиаторы и нейрогормоны
Нервные клетки управляют функциями организма с помощью химических сигнальных
Нейромедиаторы — короткоживущие вещества локального действия; они выделяются в синаптическую щель и передают сигнал соседним клеткам.
Нейрогормоны — долгоживущие вещества дальнего действия, поступающие в кровь. Однако граница между двумя группами достаточно условная, поскольку большинство медиаторов одновременно действует как гормоны.

Слайд 36Критерии, по которым вещество может быть отнесено к медиаторам.
В нервных волокнах
При раздражении нерва вещество должно выделяться и реагировать со специальным рецептором и вызывать биологическую реакцию.
Должны существовать механизмы, прекращающие действие медиатора.

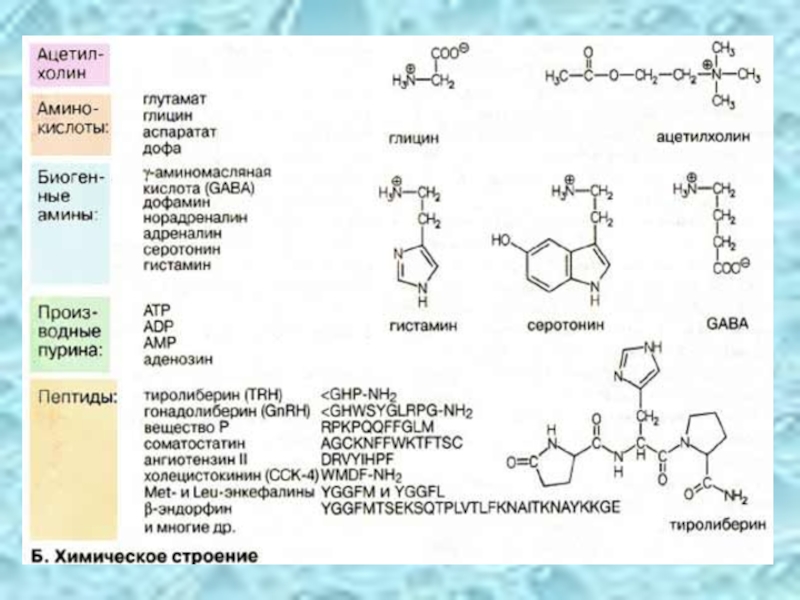
Слайд 38Химическое строение
По химическим свойствам нейромедиаторы подразделяются на несколько групп. В таблице
Наиболее известным и часто встречающимся нейромедиатором является ацетилхолин, сложный эфир холина и уксусной кислоты. К нейромедиаторам относятся некоторые аминокислоты, а также биогенные амины, образующиеся при декарбоксилировании аминокислот. Известные нейромедиаторы пуринового ряда — производные аденина. Самую большую группу образуют пептиды и белки. Небольшие пептиды часто несут на N-конце остаток глутаминовой кислоты в виде циклического пироглутамата (5-оксопролин; однобуквенный код:

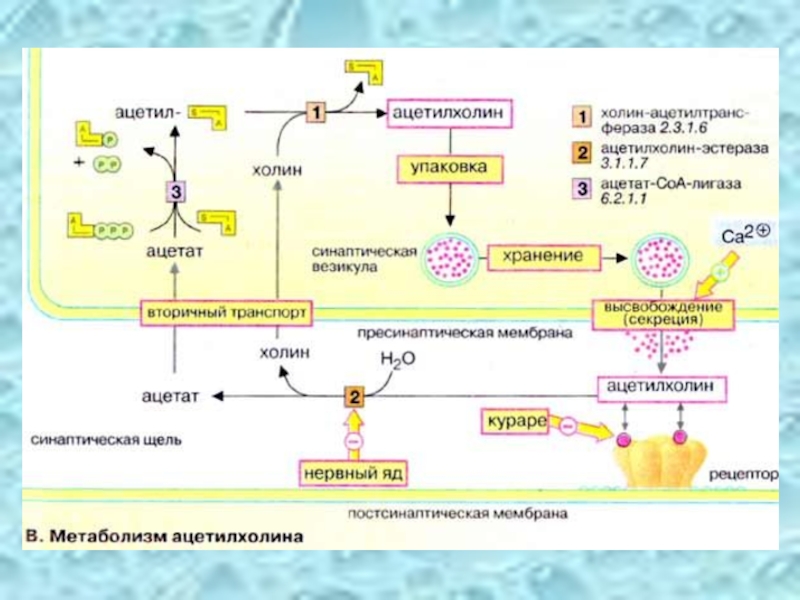
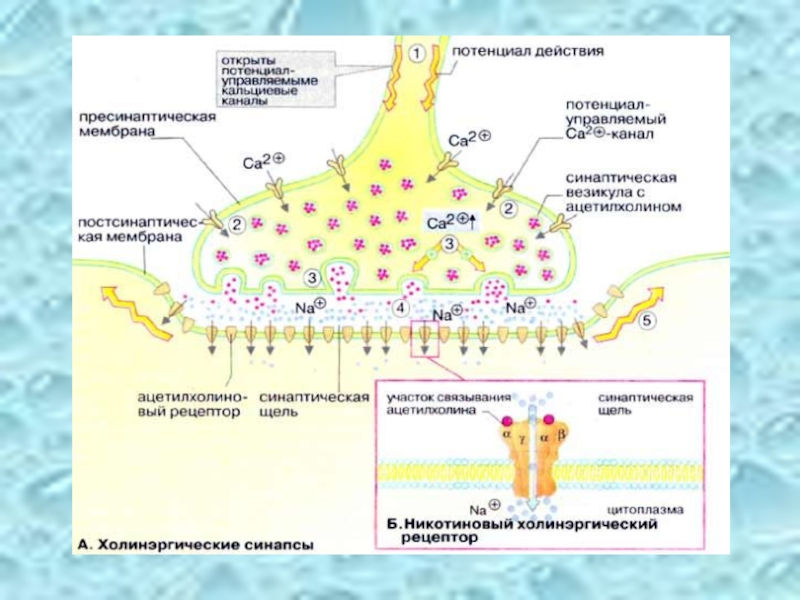
Слайд 39Ацетилхолин – сложный эфир уксусной кислоты и холина.
Деполяризация мембраны синаптических
Изменяется проницаемость мембраны – увеличивается её пропускная способность для ионов натрия.
Инактивация ацетилхолина в холинэргических синапсах:
ацетилхолин
ацетилхолин эстераза холин + уксусная кислота
Активный транспорт ацетилхолина в нейрон, где он накапливается для последующего повторного использования.

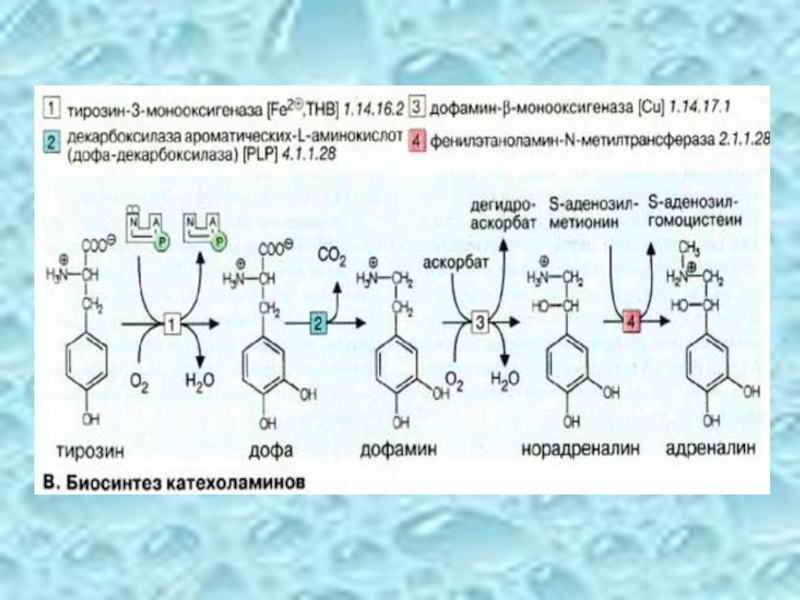
Слайд 41Норадреналин действует на адренэргические рецепторы. Повышение цАМФ приводит
Ферменты синтеза катехоламинов образуются в теле нейронов и с аксоплазматическим током транспортируются в окончания нервов.
Инактивация
МАО инактивирует норадреналин, дофамин, серотонин в пресинаптической мембране.
Норадреналин вторично поглощается симпатическими нервами

Слайд 42Дофамин, серотонин, ГАМК.
Депрессия связана с недостатком катехоламинов. Нарушение
ГАМК – медиатор торможения. При недостатке ГАМК у детей после рождения возникают судороги.
Резерпин ингибирует депонирование катехоламинов в синаптических пузырьках. Применяется для снижения артериального давления, при лечении шизофрении.
Аминазин и галоперидол блокируют дофаминовые рецепторы. При шизофрении усилена дофаминэргическая импульсация и эти лекарства целесообразны.

Слайд 43Глицин – медиатор торможения, подобный ГАМК. Функционирует в синапсах спинного мозга.
НАД нейротропен, применяется при лечении психозов. НАД специфически взаимодействует с ГАМК – бензодиазепиновым рецепторным комплексом синаптических мембран.
Нейротрубочки и нейрофиламенты – основные цитоплазматические органеллы аксона. Нейротрубочки состоят из глобулярного гликопротеида тубулина. ГТФ инициирует активацию белка. Микрофиламенты – тонкие цитоплазматические белковые нити. Особенность химического состава и мозга – присутствие в нём 2-х сильнокислых белков: S-100 содержится в глие изменение его свойств нарушает структуру нейронов и проведение нервных импульсов. Белок 14-3-2 в сером веществе содержится, перемещается из тела клетки системой медленного транспорта.


Слайд 47
В сетчатке глаза позвоночных содержатся два типа фоторецепторных клеток: палочки и
Механизм зрительного восприятия

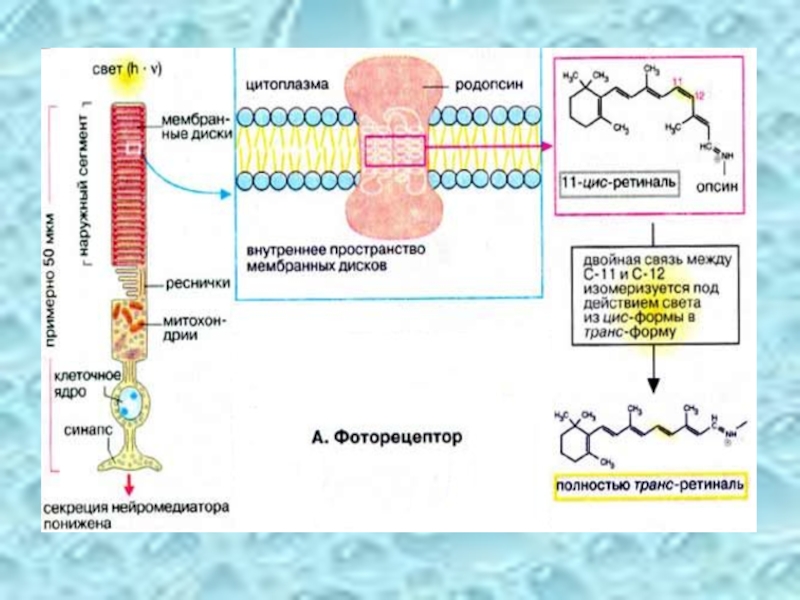
Слайд 49Фоторецептор
На рисунке схематически изображена одна из фоторецепторных клеток, палочка. Клетка состоит
Поглощение молекулой родопсина кванта света индуцирует изомеризацию 11-цис-ретиналя в полностью транс-форму. В результате этой фотохимической реакции изменяется геометрия ретиналя, а спустя 10 мс происходит аллостерический переход родопсина в его активную форму (родопсин*). Стимуляция родопсином* G-белка запускает каскад передачи сигнала, который побуждает зрительную клетку уменьшить выброс нейромедиатора (глутамата), вследствие чего биполярные нейроны, связанные со зрительными клетками, посылают измененный импульс, что воспринимается как зрительное возбуждение.

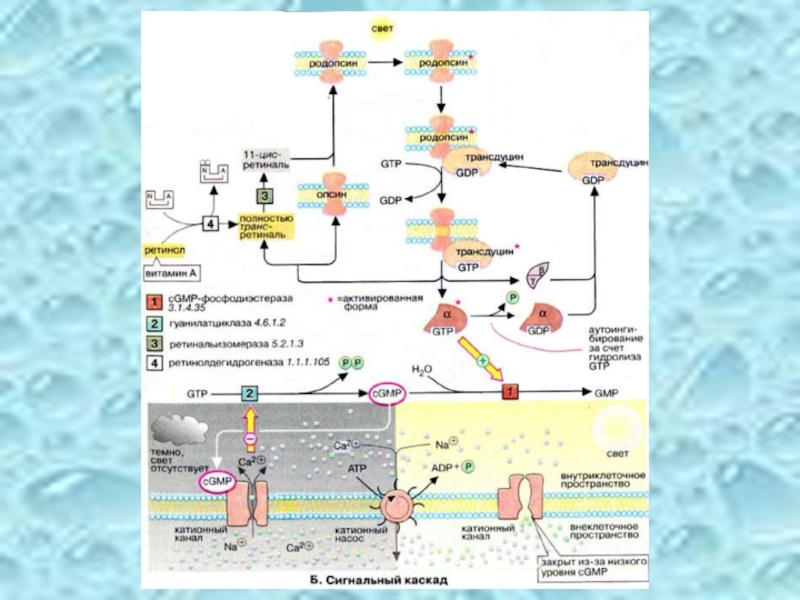
Слайд 51Сигнальный каскад
В отсутствие света концентрация цГМФ (cGMP) в колбочках поддерживается на сравнительно высоком уровне (70 мкм). Этот вторичный мессенджер постоянно синтезируется гуанилатциклазой и гидролизуется цГМФ-фосфодиэстеразой. Активация фосфодиэстеразы (при освещении родопсина) вызывает быстрое (в течение нескольких мс) падение уровня цГМФ.
Спустя короткое время α-субьединица трансдуцина инактивируется за счет медленного гидролиза связанного ГТФ и ассоциирует с комплексом β, γ-субъединиц. Родопсин* распадается на опсин и полностью транс-ретиналь, который изомеризуется в цис-ретиналь под действием изомеразы. После сборки родопсина молекула возвращается в исходное состояние.

Слайд 52В темноте (на схеме слева внизу) высокий уровень цГМФ в палочках
При освещении (на схеме справа внизу) уровень цГМФ резко падает за счет активации фосфодиэстеразы*, что приводит к перекрыванию ионных каналов. Так как ионы Na+ и Ca2+ постоянно выкачиваются из клетки, концентрация их быстро падает. Это приводит к гиперполяризации клетки и останавливает выброс нейромедиатора. Снижение концентрации ионов Ca2+ инициирует активацию гуанилатциклазы, что влечет за собой быстрый подъем уровня цГМФ настолько, что ионные каналы открываются вновь.
 высокий уровень цГМФ в палочках поддерживается благодаря активности гуанилатциклазы.")