И НАУКИ
Федеральное государственное бюджетное образовательное учреждение
высшего профессионального образования
ТЮМЕНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Институт биологии
Кафедра экологии и генетики
- Главная
- Разное
- Дизайн
- Бизнес и предпринимательство
- Аналитика
- Образование
- Развлечения
- Красота и здоровье
- Финансы
- Государство
- Путешествия
- Спорт
- Недвижимость
- Армия
- Графика
- Культурология
- Еда и кулинария
- Лингвистика
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Детские презентации
- Информатика
- История
- Литература
- Маркетинг
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Апоптоз. Морфологические проявления апоптоза презентация
Содержание
- 1. Апоптоз. Морфологические проявления апоптоза
- 2. Апоптоз — форма гибели клетки, проявляющаяся в
- 3. Исследования феномена программируемой клеточной смерти начались с
- 4. Для чего нужен апоптоз? Одной из основных
- 5. Механизм гормоны; антигены; моноклональные антитела и др.
- 8. Несмотря на разнообразие инициирующих факторов, выделяются два
- 9. Основные направления разрушительного действия каспаз на клетку:
- 10. Механизм регуляции с помощью цитокинов (путь «рецепторов
- 11. В настоящее время известно еще
- 12. Митохондриальный сигнальный путь
- 13. Раскрытие пор стимулируют следующие факторы:
- 14. Цитохром c в цитоплазме клетки участвует в формировании апоптосомы вместе
- 15. К тому же открывается доступ CARD-домена APAF-1
- 17. Эндоплазматический ретикулум и его роль в апоптозе
- 18. Другие индукторы апоптоза.
- 19. Причины апоптоза Феномен апоптоза является результатом действия различных факторов, приводящих к гибели клетки:
- 20. опухолевые заболевания аутоимунные заболевания нейродегенеративные заболевания
- 21. Заключение Апоптоз является генетически запрограммированным защитным механизмом,
- 22. Источники литературы Григорьев М. Ю. Апоптоз в
- 23. Благодарю за внимание!!!
Слайд 1Выполнил:
студент 2 курса магистратуры
Наргужина Жамила
Проверил:
к.б.н.
Тупицина Л. С.
Апоптоз
РОССИЙСКАЯ ФЕДЕРАЦИЯ
МИНИСТЕРСТВО ОБРАЗОВАНИЯ

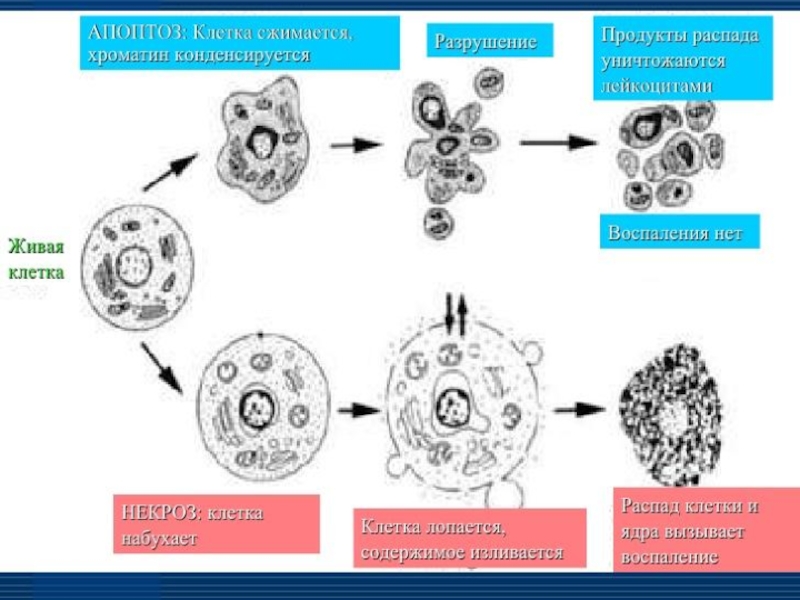
Слайд 2Апоптоз — форма гибели клетки, проявляющаяся в уменьшении ее размера, конденсации
и фрагментации хроматина, уплотнении наружной и цитоплазматических мембран без выхода содержимого клетки в окружающую среду.

Слайд 3Исследования феномена программируемой клеточной смерти начались с конца 1960-х годов. Одним
из первых в данной области был Джон Керр, который изучал гибель гепатоцитов при остром поражении печени у крыс.
В 1972 году коллектив британских учёных во главе с Дж. Керром впервые предложили использовать термин «апоптоз» для обозначения программируемой клеточной смерти.
14 марта 2000 года Джон Керр был удостоен престижной премии Пауля Эрлиха и Людвига Дармштедтера за свой вклад в исследования апоптоза.
В 1972 году коллектив британских учёных во главе с Дж. Керром впервые предложили использовать термин «апоптоз» для обозначения программируемой клеточной смерти.
14 марта 2000 года Джон Керр был удостоен престижной премии Пауля Эрлиха и Людвига Дармштедтера за свой вклад в исследования апоптоза.

Слайд 4Для чего нужен апоптоз?
Одной из основных функций апоптоза является уничтожение дефектных
клеток. В многоклеточных организмах апоптоз к тому же задействован в процессах дифференциации и морфогенеза, в поддержании клеточного гомеостаза, в обеспечении важных аспектов развития и функционирования иммунной системы. Апоптоз наблюдается у всех эукариот, начиная от одноклеточных простейших и вплоть до высших организмов.
Апоптоз во время нормального развития конечности мыши. Клетки подвергшиеся апоптозу (слева) мечены желтым. Та же конечность (справа) через один день.



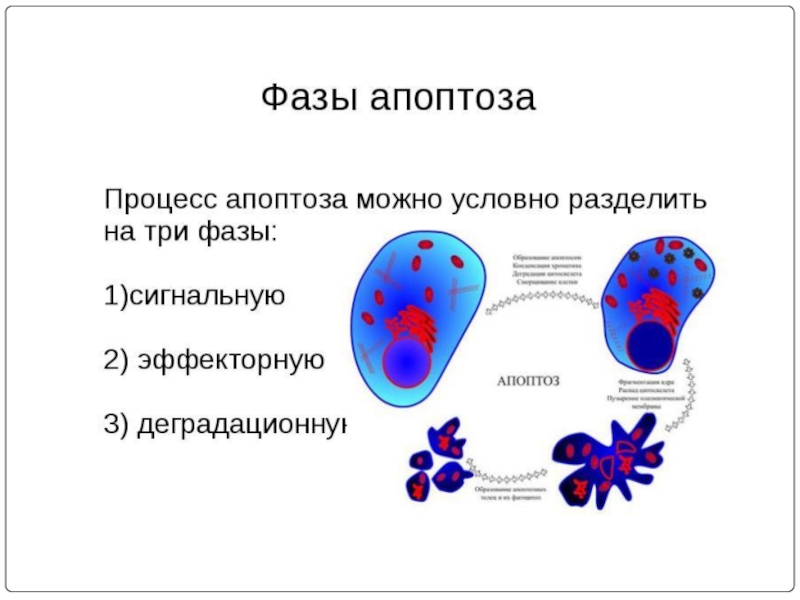
Слайд 8Несмотря на разнообразие инициирующих факторов, выделяются два основных пути трансдукции сигнала
апоптоза:
1)рецептор-зависимый сигнальный путь с участием рецепторов гибели клетки
2)митохондриальный путь.
1)рецептор-зависимый сигнальный путь с участием рецепторов гибели клетки
2)митохондриальный путь.
рецептор-зависимый сигнальный путь с")
Слайд 9Основные направления разрушительного действия каспаз на клетку:
Инактивация ингибиторов апоптотических белков:
CAD (каспазактивированная ДНК-аза) соединена с белком-ингибитором IСАD; активированные каспазы 3 и 7 разрушают белок ICAD, запуская действие ДНК-азы.
2. Прямое расщепление структурных белков клетки: ядерный ламин — белок, жестко связанный с ядерной мембраной и организующий структуру хроматина; активированная каспаза 6 расщепляет ламин, приводя к конденсации хроматина.
3. Нарушение регуляции белкового синтеза: гелсо- лин регулирует натяжение нитей актина; эффекторные каспазы расщепляют гелсолин, нарушая цитоскелет.
4. Инактивация белков, вовлеченных в репарацию ДНК, образование мРНК, репликацию ДНК.
2. Прямое расщепление структурных белков клетки: ядерный ламин — белок, жестко связанный с ядерной мембраной и организующий структуру хроматина; активированная каспаза 6 расщепляет ламин, приводя к конденсации хроматина.
3. Нарушение регуляции белкового синтеза: гелсо- лин регулирует натяжение нитей актина; эффекторные каспазы расщепляют гелсолин, нарушая цитоскелет.
4. Инактивация белков, вовлеченных в репарацию ДНК, образование мРНК, репликацию ДНК.
 соединена")
Слайд 10Механизм регуляции с помощью цитокинов (путь «рецепторов смерти»)
Цитокины — группа белков,
которые посредством связывания со специфическими рецепторами на клетках-мишенях регулируют их пролиферацию и дифференцировку.
Fas-лиганд (FasL) — цитокин семейства фактора некроза опухолей, вызывающий при взаимодействии со специфическим рецептором Fas/АРО/CD95 апоптоз клетки.
Fas-лиганд (FasL) — цитокин семейства фактора некроза опухолей, вызывающий при взаимодействии со специфическим рецептором Fas/АРО/CD95 апоптоз клетки.
Цитокины — группа белков, которые посредством связывания со")
Слайд 11 В настоящее время известно еще 4 специфических рецептора к
белкам смерти (рецепторы смерти): TNFR1; DR3 (TRAMP, Apo-3, Wsl-1); DR-4 (TRAIL-R1); DR5 (TRAIL-R2).
Помимо перечисленных соединений, в клетках существует механизм негативной регуляции этого процесса — эндогенные ингибиторы К подавляющие активность К 3 и 9 и структурные гомологи проксимальных К 8 и 10. К последним относится клеточный протеин, известный как FLIP, который способен угнетать активацию К 8 и 10
Помимо перечисленных соединений, в клетках существует механизм негативной регуляции этого процесса — эндогенные ингибиторы К подавляющие активность К 3 и 9 и структурные гомологи проксимальных К 8 и 10. К последним относится клеточный протеин, известный как FLIP, который способен угнетать активацию К 8 и 10
: TNFR1;")

Слайд 13Раскрытие пор стимулируют следующие факторы:
1)неорганический фосфат;
2)каспазы;
3)SH-реагенты;
4)истощение клеток восстановленным глутатионом
5)образование активных форм кислорода; разобщение окислительного фосфорилирования протонофорными соединениями;
6)увеличение содержания Ca в цитоплазме;
7)воздействие церамида;
8)истощение митохондриального пула АТФ .
неорганический фосфат; 2)каспазы;3)SH-реагенты;4)истощение клеток восстановленным глутатионом 5)образование активных форм кислорода;")
Слайд 14Цитохром c в цитоплазме клетки участвует в формировании апоптосомы вместе с белком APAF-1 («активирующий
фактор апоптотической протеазы-1»). Предполагается, что трансформированный APAF-1 приобретает способность связывать цитохром c.
апоптосома
. Предполагается,")
Слайд 15К тому же открывается доступ CARD-домена APAF-1 для прокаспазы-9. В итоге
происходит олигомеризация 7 субъединиц трансформированного белка APAF-1 с участием цитохрома c и прокаспазы-9. Так образуется апоптосома, активирующая каспазу-9. Зрелая каспаза-9 связывает и активирует прокаспазу-3 с образованием эффекторной каспазы-3. Высвобождающийся из межмембранного пространства митохондрий флавопротеин AIF является эффектором апоптоза, действующим независимо от каспаз.
Апоптоз

Слайд 17Эндоплазматический ретикулум и его роль в апоптозе
эндоплазматический ретикулум (ЭР) рассматривается
как участник индуцированного стрессом апоптоза
 рассматривается как участник индуцированного стрессом апоптоза")

Слайд 19Причины апоптоза
Феномен апоптоза является результатом действия различных факторов, приводящих к гибели
клетки:

Слайд 20опухолевые заболевания
аутоимунные заболевания
нейродегенеративные заболевания
Нога человека со сросшимися указательным и средним пальцами
иллюстрирует результат незавершившегося или нарушенного апоптоза на стадии эмбриогенеза.

Слайд 21Заключение
Апоптоз является генетически запрограммированным защитным механизмом, который направлен на запуск самоуничтожения
патологически измененных, мутировавших клеток (содержащих дефектные ДНК), ради сохранения целостности макроорганизма.
Как правило, борьба с дефектными клетками не ограничивается только запуском апоптоза, и протекает при активации реакций клеточного и гуморального иммунитета.
Проявлением недостаточности апоптоза служит неконтролируемое деление атипичных клеток, то есть образование и рост опухоли.
В то же время, усиленный апоптоз может приводить к раннему старению, развитию клеточной аплазии и дегенерации.
В настоящее время при разработке современных методов противоопухолевого лечения немало внимания уделяется процессам клеточной регуляции и индукции апоптоза.
Как правило, борьба с дефектными клетками не ограничивается только запуском апоптоза, и протекает при активации реакций клеточного и гуморального иммунитета.
Проявлением недостаточности апоптоза служит неконтролируемое деление атипичных клеток, то есть образование и рост опухоли.
В то же время, усиленный апоптоз может приводить к раннему старению, развитию клеточной аплазии и дегенерации.
В настоящее время при разработке современных методов противоопухолевого лечения немало внимания уделяется процессам клеточной регуляции и индукции апоптоза.

Слайд 22Источники литературы
Григорьев М. Ю. Апоптоз в норме и патологии / М.
Ю. Григорьев, Е. Н. Имянитов, К. П. Хансон // Медицинский академический журнал. — 2003. — Т. 3, № 3. — С. 3—11.
Владимирская Е. Б. Механизмы апоптотической смерти клеток / Е. Б. Владимирская // Гематология и транс- фузиология. — 2002. — Т. 47, № 2. — С. 35—40.
Залесский В. Н. Перспективы патогенетически обос- нованного применения модуляторов апоптоза в качестве нейро-, кардио-, гепато- и нефропротекторов (обзор лите- ратуры) / В. Н. Залесский, А. А. Фильченков // http://www. medved.kiev.ua/arhiv_mg/st_2001/01_4_16.htm
Робинсон М. В. Апоптоз клеток иммунной системы / М. В. Робинсон, В. А. Труфакин // Успехи современной биологии. — 1991. — Т. 3, вып. 2. — С. 246—259
Владимирская Е. Б. Механизмы апоптотической смерти клеток / Е. Б. Владимирская // Гематология и транс- фузиология. — 2002. — Т. 47, № 2. — С. 35—40.
Залесский В. Н. Перспективы патогенетически обос- нованного применения модуляторов апоптоза в качестве нейро-, кардио-, гепато- и нефропротекторов (обзор лите- ратуры) / В. Н. Залесский, А. А. Фильченков // http://www. medved.kiev.ua/arhiv_mg/st_2001/01_4_16.htm
Робинсон М. В. Апоптоз клеток иммунной системы / М. В. Робинсон, В. А. Труфакин // Успехи современной биологии. — 1991. — Т. 3, вып. 2. — С. 246—259







